PRODUCT CENTER
特色產(chǎn)品當(dāng)前位置:首頁 > 特色產(chǎn)品
新血管的形成(angiogenesis)是腫瘤生長轉(zhuǎn)移和傳播過程中的一種基本活動(dòng)。因此,在癌癥研究領(lǐng)域,人們對(duì)研究腫瘤血管生成的分子機(jī)制十分感興趣。而血管內(nèi)皮生長因子(vascular endothelial growth factor,VEGF)路徑是這一過程的關(guān)鍵調(diào)節(jié)者。
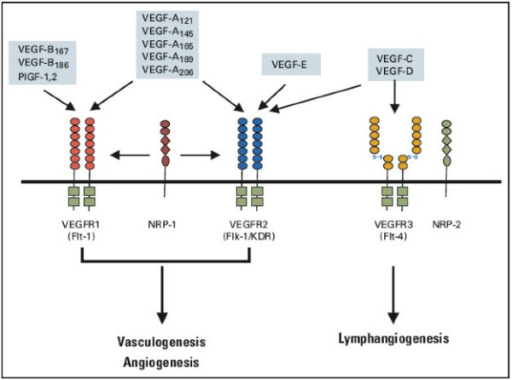
VEGF(vascular endothelial growth factor,血管內(nèi)皮生長因子)/VEGFR(vascular endothelial growth factor receptor,血管內(nèi)皮生長因子受體)軸由多重配基和受體質(zhì)量疊加交錯(cuò)組成,并且受體與配基結(jié)合具有專一性,在不同的細(xì)胞中具有不同的細(xì)胞類型表達(dá)和功能,啟動(dòng)VEGFR信號(hào)通路,觸發(fā)了一個(gè)網(wǎng)狀的信號(hào)過程,從而促進(jìn)血管內(nèi)皮細(xì)胞生長、轉(zhuǎn)移和存活。VEGF/VEGFR也是目前研究最多的一條信號(hào)通路,并取得了顯著的成果。

作為專業(yè)的生命科學(xué)醫(yī)藥原料和解決方案供應(yīng)商,艾美捷科技為您推薦來自德國,Relia Tech榮譽(yù)出品:活性VEGF,VEGFR重組蛋白
| 貨號(hào) | 中文名稱 | 來源 | 規(guī)格 | 氨基酸(a.a) | Uniprot |
| RET-M30-032 | 重組蛋白, 小鼠, VEGF120 | 大腸桿菌 | 20ug | 120 | Q00731 |
| RET-R20-064 | 重組蛋白, 大鼠, VEGF120 | 大腸桿菌 | 20ug | 120 | P16612 |
| RET-300-072 | 重組蛋白, 人, VEGF121 | 大腸桿菌 | 20ug | 121 | P15692 |
| RET-300-032 | 重組蛋白, 人, VEGF121 | 昆蟲細(xì)胞 | 20ug | 121 | P15692 |
| RET-M30-034 | 重組蛋白, 小鼠, VEGF144 | 大腸桿菌 | 20ug | 144 | Q00731 |
| RET-300-033S-E | 重組蛋白, 人, VEGF145 | 大腸桿菌 | 2ug | 145 | P15692 |
| RET-300-034-E | 重組蛋白, 人, VEGF145 | 大腸桿菌 | 20ug | 145 | P15692 |
| RET-M30-004 | 重組蛋白, 小鼠, VEGF164 | 大腸桿菌 | 20ug | 164 | Q00731 |
| RET-M30-002 | 重組蛋白, 小鼠, VEGF164 | 昆蟲細(xì)胞 | 20ug | 164 | Q00731 |
| RET-R20-068 | 重組蛋白, 大鼠, VEGF164 | 大腸桿菌 | 20ug | 164 | P16612 |
| RET-300-076 | 重組蛋白, 人, VEGF165 | 大腸桿菌 | 20ug | 164 | P15692 |
| RET-300-036 | 重組蛋白, 人, VEGF165 | 昆蟲細(xì)胞 | 20ug | 165 | P15692 |
| RET-300-082 | 重組蛋白, 人, VEGF165b | 大腸桿菌 | 20ug | 164 | P15692-8 |
| RET-300-065Bi | 重組蛋白, 人, VEGF165-生物素 | 大腸桿菌 | 10ug | 165 | P15692 |
| RET-300-066Bi | 重組蛋白, 人, VEGF165-生物素 | 大腸桿菌 | 25ug | 165 | P15692 |
| RET-M30-095 | 重組蛋白, 小鼠, VEGF188 | 大腸桿菌 | 20ug | 188 | Q00731 |
| RET-R20-070 | 重組蛋白, 大鼠, VEGF188 | 大腸桿菌 | 20ug | 188 | P16612 |
| RET-300-095 | 重組蛋白, 人, VEGF189 | 大腸桿菌 | 20ug | 189 | P15692 |
| RET-300-099 | 重組蛋白, 人, VEGF206 | 大腸桿菌 | 20ug | 206 | P15692 |
| RET-300-080 | 重組蛋白, 人, VEGF-B167 | 大腸桿菌 | 20ug | 167 | P49765 |
| RET-300-079 | 重組蛋白, 人, VEGF-C | 昆蟲細(xì)胞 | 20ug | 121 | P49767 |
| RET-R20-015 | 重組蛋白, 大鼠, VEGF-C | 昆蟲細(xì)胞 | 20ug | 127 | O35757 |
| RET-R20-017 | 重組蛋白, 大鼠, VEGF-C152S | 昆蟲細(xì)胞 | 20ug | 127 | O35757 |
| RET-300-045 | 重組蛋白, 羊口瘡病毒, VEGF-E | 大腸桿菌 | 20ug | 132 | Q9YMF3 |
| RET-300-046 | 重組蛋白, 羊口瘡病毒, VEGF-E, 肝素結(jié)合域 | 昆蟲細(xì)胞 | 20ug | 154 | Q9YMF3 |
| RET-300-097 | 重組蛋白, 蛇, VEGF-F (矛頭蝮) | 大腸桿菌 | 20ug | 124 | Q90X24 |
| RET-S01-016 | 重組蛋白, 人, VEGFR-1/Flt-1 (D3), 可溶性蛋白 | 昆蟲細(xì)胞 | 20ug | 327 | P17948 |
| RET-S01-080 | 重組蛋白, 人, VEGFR-1/Flt-1 (D3)-His, 可溶性蛋白 | 昆蟲細(xì)胞 | 50ug | 327 | P17948 |
| RET-S01-014 | 重組蛋白, 人, VEGFR-1/Flt-1 (D4), 可溶性蛋白 | 昆蟲細(xì)胞 | 20ug | 457 | P17948 |
| RET-S01-012 | 重組蛋白, 人, VEGFR-1/Flt-1 (D5), 可溶性蛋白 | 昆蟲細(xì)胞 | 20ug | 536 | P17948 |
| RET-S01-010 | 重組蛋白, 人, VEGFR-1/Flt-1(天然), 可溶性蛋白 | 昆蟲細(xì)胞 | 20ug | 661 | P17948 |
| RET-S01-072 | 重組蛋白, 人, VEGFR1-14/Flt1-14, 可溶性蛋白 | 昆蟲細(xì)胞 | 20ug | 707 | P17948-3 |
| RET-SFC-006 | 重組蛋白, 人, VEGFR-1/Flt-1(D7)-Fc 嵌合體, 可溶性蛋白 | 昆蟲細(xì)胞 | 50ug | 954 | P17948 |
| RET-SFC-M06 | 重組蛋白, 小鼠, VEGFR-1/Flt-1(D7)-Fc 嵌合體, 可溶性蛋白 | 昆蟲細(xì)胞 | 50ug | 965 | P17948 |
| RET-S01-002 | 重組蛋白, 人, VEGFR-2/KDR (D7), 可溶性蛋白 | 昆蟲細(xì)胞 | 50ug | 738 | P35968 |
| RET-S01-004 | 重組蛋白, 人, VEGFR-2/KDR(天然), 可溶性蛋白 | 昆蟲細(xì)胞 | 20ug | 659 | P35968 |
| RET-SFC-008 | 重組蛋白, 人, VEGFR-2/KDR-Fc 嵌合體, 可溶性蛋白 | 昆蟲細(xì)胞 | 50ug | 968 | P35968 |
| RET-S01-M04 | 重組蛋白, 小鼠, VEGFR-2/Flk-1(天然), 可溶性蛋白 | 昆蟲細(xì)胞 | 20ug | 654 | P35918 |
| RET-S01-018 | 重組蛋白, 人, VEGFR-3/FLT-4, 可溶性蛋白 | 昆蟲細(xì)胞 | 50ug | 761 | P35916 |
| RET-SFC-010 | 重組蛋白, 人, VEGFR-3/FLT-4/Fc 嵌合體, 可溶性蛋白 | 昆蟲細(xì)胞 | 50ug | 979 | P35916 |
* 以上蛋白產(chǎn)品,均為凍干粉形式.* 蛋白分子量與序列詳情,歡迎垂詢艾美捷客服.
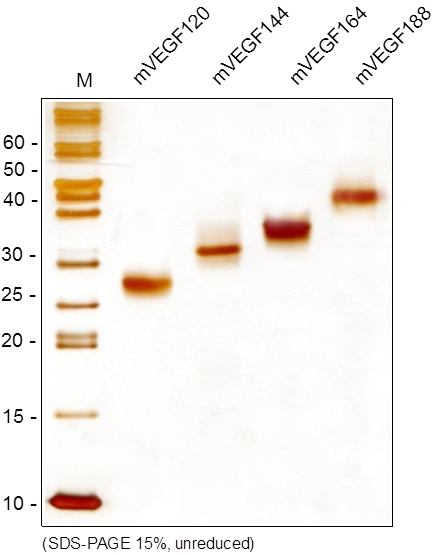 | 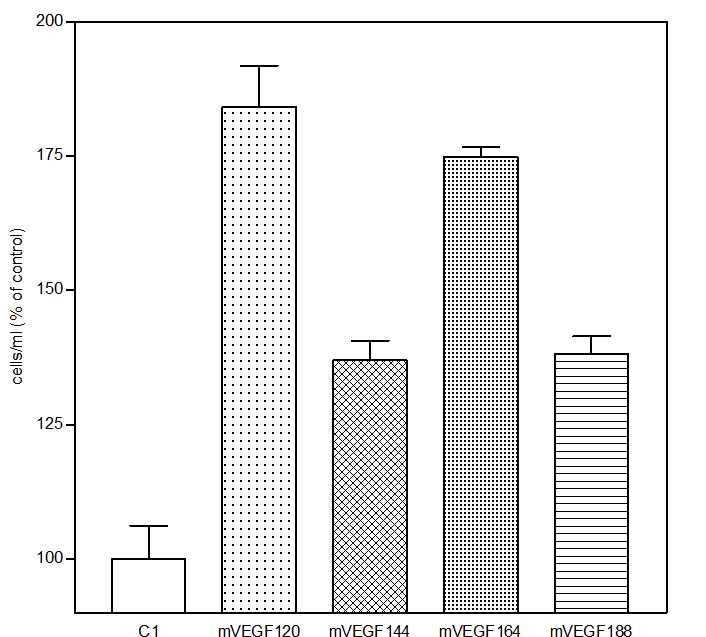 |
| VEGF蛋白 SDS-PAGE | VEGF蛋白刺激HUVEC增殖 |
【VEGF/VEGFR產(chǎn)品發(fā)表文章】
Cudmore, M., Hewett, P., Ahmad, S. et al. The role of heterodimerization between VEGFR-1 and VEGFR-2 in the regulation of endothelial cell homeostasis. Nat Commun 3, 972 (2012). https://doi.org/10.1038/ncomms1977
Teichert, M., Milde, L., Holm, A. et al. Pericyte-expressed Tie2 controls angiogenesis and vessel maturation. Nat Commun 8, 16106 (2017). https://doi.org/10.1038/ncomms16106
SUCNR1 Is Expressed in Human Placenta and Mediates Angiogenesis: Significance in Gestational Diabetes. R. Atallah et al., Int J Mol Sci. 2021 Nov; 22(21): 12048.
Transforming growth factor‐β1 signalling triggers vascular endothelial growth factor resistance and monocyte dysfunction in type 2 diabetes mellitus. L.‐M. Makowski et al., J Cell Mol Med. 2021 Jun; 25(11): 5316–5325.
Outgrowth, proliferation, viability, angiogenesis and phenotype of primary human endothelial cells in different purchasable endothelial culture media: feed wisely. B. Leopold et al., Histochem Cell Biol. 2019; 152(5): 377–390.
Apelin+ Endothelial Niche Cells Control Hematopoiesis and Mediate Vascular Regeneration after Myeloablative Injury. Qi Chen et al., Cell Stem Cell. 2019 Dec 5; 25(6): 768–783.e6.
BMP-2 induces human mononuclear cell chemotaxis and adhesion and modulates monocyte-to-macrophage differentiation. E. Pardali et al., J Cell Mol Med. 2018 Nov;22(11):5429-5438.
Hypoxia Impairs Initial Outgrowth of Endothelial Colony Forming Cells and Reduces Their Proliferative and Sprouting Potential. Tasev D et al., Front Med (Lausanne). 2018 Dec 20;5:356.
Blood Outgrowth and Proliferation of Endothelial Colony Forming Cells are Related to Markers of Disease Severity in Patients with Pulmonary Arterial Hypertension. Smits J et al., Int J Mol Sci. 2018 Nov 27;19(12). pii: E3763.
Pericytes regulate VEGF-induced endothelial sprouting through VEGFR1. H. M. Eilken et al., Nat Commun. 2017; 8: 1574.
CD34 expression modulates tube-forming capacity and barrier properties of peripheral blood-derived endothelial colony-forming cells (ECFCs). D. Tasev et al., Angiogenesis. 2016; 19: 325–338.
Endothelial Cells Derived from Non-malignant Tissues Are of Limited Value as Models for Brain Tumor Vasculature. Lohr J et al., Anticancer Res. 2015 May;35(5):2681-90.
Long-Term Expansion in Platelet Lysate Increases Growth of Peripheral Blood-Derived Endothelial-Colony Forming Cells and Their Growth Factor-Induced Sprouting Capacity. D. Tasev et al., PLoS One. 2015; 10(6): e0129935.
The inhibition of tyrosine kinase receptor signalling in leiomyosarcoma cells using the small molecule kinase inhibitor PTK787/ZK222584 (Vatalanib?). A. K.A. Gaumann et al., Int J Oncol. 2014 Dec; 45(6): 2267–2277.
Improved Anchorage of Ti6Al4V Orthopaedic Bone Implants through Oligonucleotide Mediated Immobilization of BMP-2 in Osteoporotic Rats. J. V. W?lfle et al., PLoS One. 2014; 9(1): e86151.
Spatial regulation of VEGF receptor endocytosis in angiogenesis. M. Nakayama et al., Nat Cell Biol.2 13 Mar; 15(3): 249–260.
Fbxw7 Controls Angiogenesis by Regulating Endothelial Notch Activity. N. Izumi et al., PLoS One. 2012; 7(7): e41116.
The Antiangiogenic 16K Prolactin Impairs Functional Tumor Neovascularization by Inhibiting Vessel Maturation. Ngoc-Quynh-Nhu Nguyen et al., PLoS One. 2011; 6(11): e27318.
MicroRNA-21 Exhibits Antiangiogenic Function by Targeting RhoB Expression in Endothelial Cells. C. Sabatel et al., PLoS One. 2011; 6(2): e16979.
Autocrine activity of soluble Flt-1 controls endothelial cell function and angiogenesis. S. Ahmad et al., Vasc Cell. 2011; 3: 15.
Feed-forward Signaling by Membrane-bound Ligand Receptor Circuit: THE CASE OF NOTCH DELTA-LIKE 4 LIGAND IN ENDOTHELIAL CELLS. V. Caolo et al., J Biol Chem. 2010 Dec 24; 285(52): 40681–40689.
Impaired Collateral Recruitment and Outward Remodeling in Experimental Diabetes. J. M. van Golde et al., Diabetes. 2008 Oct; 57(10): 2818–2823.
Elevated placental soluble vascular endothelial growth factor receptor-1 inhibits angiogenesis in preeclampsia. S. Ahmad and A. Ahmed Circ Res. 2004 Oct 29;95(9):884-91.
Collagen type 1 retards tube formation by human microvascular endothelial cells in a fibrin matrix. Kroon, M.E. et al., Angiogenesis (2002) 5: 257.
德國重組蛋白品牌 ReliaTech ,ISO9001認(rèn)證,低內(nèi)毒素,90%以上的產(chǎn)品均有現(xiàn)貨,每3周發(fā)貨 ,快行業(yè)2周!
熱銷:FGF2、a-mLYVE1、VEGF等活性重組蛋白【已發(fā)表文章超1000篇】,2800+細(xì)胞培養(yǎng)級(jí)低內(nèi)毒素重組蛋白和抗體,專注受體與配體研究,適合用于VEGF靶向藥物研發(fā)、生物制藥原材料。
http://m.zihai029.com/brand/ReliaTech.shtml
-----------------------------------------------------------------------
以上所有產(chǎn)品,均僅限于科研用途。不得用于醫(yī)療。
-----------------------------------------------------------------------
作為一家具有高端的技術(shù)實(shí)力、先進(jìn)的經(jīng)營管理水平和完善的市場銷售體系的生物高科技企業(yè),總部位于武漢光谷高新技術(shù)開發(fā)區(qū),服務(wù)面向全國。艾美捷科技是集進(jìn)口試劑、實(shí)驗(yàn)室耗材銷售、技術(shù)服務(wù)與合約開發(fā)為一體的專業(yè)化高科技公司,為用戶提供專業(yè)的前沿資訊、完備的產(chǎn)品、整合的解決方案,及優(yōu)質(zhì)的物流服務(wù)。
艾美捷科技與國內(nèi)外優(yōu)秀的生物試劑供應(yīng)商優(yōu)保持著密切的合作關(guān)系,目前已成為眾多國際知名品牌的中國總代理或一級(jí)代理,主要包括:AAT Bioquest、Abbexa、Abnova、Agrisera、Atlas Antibodies、BellBrook、Biomatik、Biosensis、BioVendor、CalBioreagents、Cayman Chem、Cell Biolabs、Columbia Biosciences、Crystal Chem、Cytoskeleton、DIAsource、Duchefa、Ebba Biotech、Echelon Biosciences、ECM Biosciences、Enzo Life Sciences、Epigentek、Equitech-Bio、FabGennix、G-Biosciences、GeneCopoeia、Hycult Biotech、ichorbio、Icosagen、Immundiagnostik、ImmunoReagents、IQ Products、Jackson、LC Labs、LifeSensors、Lumiprobe、Mabtech、Matreya、Medkoo Biosciences、MyBioSource、Nanoprobe、Norgen Biotek、ProSci、ProSpec、ReliaTech、Rockland、SouthernBiotech、StressMarq、SySy、TRC、US Biological、Worthington 等,可以在短時(shí)間內(nèi)為用戶提供專業(yè)的前沿資訊、完備的產(chǎn)品及物流服務(wù)。

產(chǎn)品訂購:sales@amyjet.com
郵政編碼:430070
公司地址:武漢市洪山區(qū)光谷大道35號(hào)
光谷總部國際二期時(shí)代1棟13樓
提示:本公司所有產(chǎn)品僅供科研使用,不用于臨床診斷。
版權(quán)所有:艾美捷科技有限公司 鄂ICP備10204150號(hào)-1 鄂公網(wǎng)安備:42018502004523號(hào)
第二類醫(yī)療器械經(jīng)營備案憑證:鄂漢藥監(jiān)械經(jīng)營備20234324號(hào)
友情鏈接:亞科因生物每日生物評(píng)論

微信掃碼在線客服