AGENT BRAND
品牌專題補(bǔ)體系統(tǒng)是先天免疫的一個(gè)重要組成部分,其高度依賴C3的激活過程。作為補(bǔ)體系統(tǒng)的第三個(gè)組分,C3對于免疫系統(tǒng)中經(jīng)典途徑、旁路途徑和凝集素途徑的有效運(yùn)作至關(guān)重要。C3的激活在免疫防御機(jī)制中發(fā)揮著基礎(chǔ)性作用,因此,對C3及其片段的理解和檢測至關(guān)重要。在免疫學(xué)研究領(lǐng)域,對C3檢測的關(guān)注度日益增加,特別是研究抗體如何與激活的C3相互作用。這種對C3檢測的關(guān)注對于推進(jìn)我們對免疫學(xué)的認(rèn)識以及開發(fā)新的治療策略具有關(guān)鍵意義。
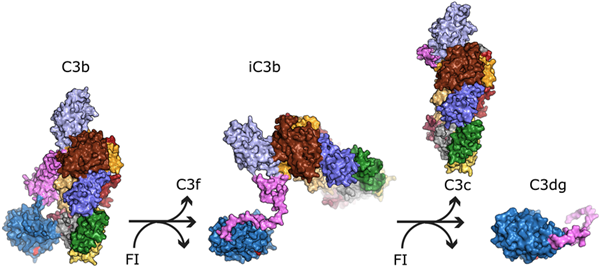
C3通過不同的結(jié)合位點(diǎn)與大量的補(bǔ)體因子(例如蛋白酶、受體和調(diào)節(jié)因子)以及非補(bǔ)體蛋白(例如病毒和細(xì)菌蛋白)相互作用。C3的功能通過順序蛋白水解切割誘導(dǎo)的構(gòu)象變化來調(diào)節(jié)。成熟C3的裂解由酶復(fù)合物(即轉(zhuǎn)化酶)介導(dǎo),生成過敏毒素C3a(9 kDa)和主要片段C3b(177 kDa)。這一步驟暴露了一個(gè)隱藏的硫酯基和C3b中多個(gè)用于與補(bǔ)體蛋白相互作用的隱匿結(jié)合位點(diǎn)。新生的C3b能夠通過暴露的硫酯基與細(xì)胞和其他靶標(biāo)表面共價(jià)結(jié)合。通過表面結(jié)合的C3b與酶原因子B結(jié)合,形成旁路途徑的C3轉(zhuǎn)化酶C3bBb,從而實(shí)現(xiàn)補(bǔ)體活性的放大。但是這種轉(zhuǎn)化酶的半衰期很短。因子I在與可溶性或膜結(jié)合的輔因子共同作用下,對C3b的α鏈進(jìn)行額外的裂解(生成iC3b),防止進(jìn)一步的轉(zhuǎn)化酶形成,并顯著改變該蛋白的功能。前兩次裂解釋放出C3f(2 kDa),第三次裂解使與靶標(biāo)結(jié)合的C3dg(40 kDa)片段釋放出C3c(135 kDa)。
補(bǔ)體系統(tǒng)是免疫系統(tǒng)的重要組成部分,而C3抗體在免疫學(xué)研究中至關(guān)重要。然而,面對眾多的選擇,選擇正確的C3抗體對于確保您研究的成功至關(guān)重要。作為補(bǔ)體領(lǐng)域的研究專家,Hycult Biotech可提供高質(zhì)量的C3及其片段抗體,這些抗體經(jīng)過嚴(yán)格測試,適用于多種研究用途。
| 貨號 | 名稱 | 特異性 |
| HM2168 | Activated C3 抗體 | 可特異性識別C3b、iC3b和C3c裂解片段上的新表位,這些表位在C3激活過程中因蛋白水解切割而暴露出來,可用于檢測補(bǔ)體級聯(lián)反應(yīng)的激活和C3的裂解。 |
| HM2257 | Activated C3抗體 | 可特異性識別C3b、iC3b和C3dg上的新表位,該表位在天然C3中不存在 :這類抗體針對的是C3激活后產(chǎn)生的特定表位,這些表位在天然未激活的C3上不存在,而在C3b、iC3b和C3dg等裂解片段上出現(xiàn),有助于研究補(bǔ)體激活的不同階段。 |
| HM2377 | C3 (beta-chain)抗體 | 識別補(bǔ)體C3的β鏈 :補(bǔ)體C3的β鏈(71 kDa)除了存在于C3中,還存在于激活產(chǎn)物C3b、iC3b和C3c中,一些抗體能夠特異性識別該β鏈,可用于檢測這些補(bǔ)體成分的存在。 |
| HM2075 | C3/C3a抗體 | 識別補(bǔ)體蛋白C3a的C末端 :C3a是C3激活過程中產(chǎn)生的過敏毒素,某些抗體能夠特異性識別C3a的C末端,可用于檢測C3a的產(chǎn)生和釋放。 |
| HM2073 | C3/C3a抗體 | 與C3a上的表位反應(yīng) :這類抗體可與C3a上的特定表位發(fā)生反應(yīng),不僅能與完整的C3反應(yīng),還能與C3a反應(yīng),可用于研究C3激活過程中C3a的生成及其生物學(xué)功能。 |
| HM2072 | C3/C3b抗體 | 識別位于C3α鏈C末端360個(gè)氨基酸上的表位 :該表位存在于C3的α鏈上,因此能夠識別C3b、iC3b、C3c以及完整的C3,可用于檢測這些補(bǔ)體成分在不同生理和病理過程中的表達(dá)和作用。 |
| HM2389 | C3/C3b/iC3b (alpha-chain)抗體 | 識別補(bǔ)體C3、C3b和iC3b :一些抗體在西方印跡和免疫分析中都能識別補(bǔ)體C3、C3b和iC3b,對C3c和C3d也有一定程度的識別,但僅限于免疫分析。其特異性表位位于C3α鏈的MG8-C345C結(jié)構(gòu)域,可用于研究C3及其裂解片段在補(bǔ)體激活過程中的作用。 |
| HM2401 | C3/C3b/iC3b/C3c抗體 | 識別補(bǔ)體C3、C3b、iC3b和C3c :這類抗體的識別域被確定為MG2-MG6,能夠阻斷旁路途徑中C3轉(zhuǎn)化酶的形成,可用于研究補(bǔ)體激活的調(diào)節(jié)機(jī)制以及相關(guān)疾病的發(fā)病機(jī)制。 |
| HM2074 | C3a/C3a des Arg抗體 | 識別人C3a/C3a-desArg上的新表位 :C3a-desArg是C3a的代謝產(chǎn)物,某些抗體能夠特異性識別C3激活過程中產(chǎn)生的C3a和C3a-desArg上的新表位,可用于檢測C3a的生成及其在炎癥反應(yīng)中的作用。 |
| HM2195 | C3aR抗體 | 與人C3a受體反應(yīng) :C3a受體是C3a發(fā)揮作用的重要靶點(diǎn),一些抗體能夠特異性識別并結(jié)合人C3a受體,可用于研究C3a與其受體的相互作用及其在細(xì)胞信號轉(zhuǎn)導(dǎo)中的作用。 |
| HM2286 | C3b/iC3b抗體 | 識別人補(bǔ)體C3b/iC3b并阻斷旁路途徑的激活 :這類抗體能夠特異性識別C3b/iC3b,并通過阻斷旁路途徑的激活來抑制補(bǔ)體系統(tǒng)的過度激活,可用于研究補(bǔ)體激活的調(diào)節(jié)機(jī)制以及相關(guān)疾病的治療策略。 |
| HM2285 | C3b/iC3b抗體 | 識別C3b/iC3b并抑制經(jīng)典途徑 :一些抗體能夠特異性結(jié)合人C3以及其裂解產(chǎn)物C3b和iC3b,從而抑制經(jīng)典途徑的激活,可用于研究補(bǔ)體激活的調(diào)節(jié)機(jī)制以及相關(guān)疾病的發(fā)病機(jī)制。 |
| HM2287 | C3b/iC3b/C3d抗體 | 識別補(bǔ)體因子C3b/iC3b/C3d :這類抗體能夠特異性結(jié)合人C3以及其裂解產(chǎn)物C3b、iC3b和C3dg,可用于研究補(bǔ)體激活過程中C3及其裂解片段的生物學(xué)功能以及它們在不同生理和病理過程中的作用。 |
| HM2394 | C3b/iC3b/C3c抗體 | 識別補(bǔ)體C3、C3b、iC3b和C3c :其識別域被確定為MG4/5,能夠阻斷旁路途徑中C3轉(zhuǎn)化酶對C3的激活,可用于研究補(bǔ)體激活的調(diào)節(jié)機(jī)制以及相關(guān)疾病的發(fā)病機(jī)制。 |
| HM2395 | iC3b/C3dg/C3d抗體 | 識別補(bǔ)體iC3b、C3dg和C3d :其識別域被確定為CUB-TED,能夠阻斷旁路途徑中C3轉(zhuǎn)化酶對C3的激活,可用于研究補(bǔ)體激活的調(diào)節(jié)機(jī)制以及相關(guān)疾病的發(fā)病機(jī)制。 |
| HM2200 | C3c抗體 | 識別裂解的人C3片段C3c :這類抗體能夠識別C3c上的構(gòu)象表位,以及C3b和iC3b上的相應(yīng)表位,可用于檢測C3激活過程中C3c的生成及其在補(bǔ)體激活過程中的作用。 |
| HM2318 | C3c抗體 | 識別僅存在于人補(bǔ)體C3c上的獨(dú)特表位 :某些抗體能夠特異性識別僅存在于C3c上的獨(dú)特表位,可用于檢測C3c的存在以及研究其在補(bǔ)體激活過程中的作用。 |
| HM2422 | C3d抗體 | 識別補(bǔ)體C3dg上的新表位 :這類抗體能夠特異性識別C3dg上的新表位,但不識別C3或(i)C3b,可用于檢測C3dg的生成以及研究其在補(bǔ)體激活過程中的作用。 |
| HM2198 | C3d抗體 | 與C3d上的線性決定簇反應(yīng) :C3d是C3激活過程中的一個(gè)裂解片段,某些抗體能夠特異性識別C3d上的線性決定簇,該決定簇也存在于C3、C3b、iC3b、C3dg和C3d上,可用于檢測這些補(bǔ)體成分的存在以及研究它們在補(bǔ)體激活過程中的作用。 |
| HM2199 | C3g抗體 | 與iC3、iC3b、C3dg和C3g上的新抗原反應(yīng) :這類抗體能夠特異性識別iC3、iC3b、C3dg和C3g上的新抗原,可用于研究補(bǔ)體激活過程中這些片段的生成及其在不同生理和病理過程中的作用。 |
產(chǎn)品訂購:sales@amyjet.com
郵政編碼:430070
公司地址:武漢市洪山區(qū)光谷大道35號
光谷總部國際二期時(shí)代1棟13樓
提示:本公司所有產(chǎn)品僅供科研使用,不用于臨床診斷。
版權(quán)所有:艾美捷科技有限公司 鄂ICP備10204150號-1 鄂公網(wǎng)安備:42018502004523號
第二類醫(yī)療器械經(jīng)營備案憑證:鄂漢藥監(jiān)械經(jīng)營備20234324號

微信掃碼在線客服