AGENT BRAND
品牌專題谷氨酸是大腦中最重要的興奮性神經(jīng)遞質(zhì)。多種離子型和代謝型受體介導(dǎo)其向神經(jīng)元傳遞的興奮信號(hào)。然而,在信號(hào)調(diào)控中,"關(guān)閉"信號(hào)與"開(kāi)啟"信號(hào)同等重要。興奮性氨基酸轉(zhuǎn)運(yùn)體(EAATs)家族通過(guò)將谷氨酸回收至神經(jīng)元和星形膠質(zhì)細(xì)胞,快速清除突觸間隙及周?chē)?xì)胞外空間中的谷氨酸,從而終止其興奮信號(hào)。EAATs通過(guò)以下機(jī)制實(shí)現(xiàn)調(diào)控:
1. 維持谷氨酸信號(hào)的時(shí)間保真度:通過(guò)清除突觸間隙中的谷氨酸,防止初始信號(hào)傳遞后突觸后受體被過(guò)度激活,從而保障后續(xù)信號(hào)的準(zhǔn)確檢測(cè)。
2. 維持信號(hào)的空間保真度:防止谷氨酸從其釋放的突觸"逃逸"至非目標(biāo)區(qū)域,避免意外激活突觸外或非目標(biāo)突觸的谷氨酸受體。
3. 防止神經(jīng)毒性:細(xì)胞外谷氨酸過(guò)量會(huì)通過(guò)過(guò)度激活谷氨酸受體引發(fā)神經(jīng)毒性級(jí)聯(lián)反應(yīng),最終導(dǎo)致神經(jīng)元功能喪失和細(xì)胞死亡(稱為興奮性毒性)。
谷氨酸與陰離子的轉(zhuǎn)運(yùn)機(jī)制
EAATs作為同向轉(zhuǎn)運(yùn)體,通過(guò)以下方式完成轉(zhuǎn)運(yùn):每轉(zhuǎn)運(yùn)一個(gè)谷氨酸分子(天冬氨酸也可被轉(zhuǎn)運(yùn))進(jìn)入細(xì)胞時(shí),同步將1個(gè)K+離子運(yùn)出細(xì)胞,并攜帶3個(gè)Na+和1個(gè)H+進(jìn)入細(xì)胞(Alleva et al., 2022)。這種轉(zhuǎn)運(yùn)依賴于鈉離子的電化學(xué)梯度,并通過(guò)細(xì)胞膜上的EAAT同源或異源三聚體實(shí)現(xiàn)(Kovermann et al., 2022)。值得注意的是,EAATs不僅是谷氨酸轉(zhuǎn)運(yùn)體,還能在谷氨酸轉(zhuǎn)運(yùn)周期中作為氯離子通道(Cl+)開(kāi)放(Otis & Jahr, 1998)。
功能擴(kuò)展
-星形膠質(zhì)細(xì)胞的EAAT2(GLT-1)承擔(dān)了約90%的谷氨酸清除任務(wù),其功能缺陷與阿爾茨海默病、肌萎縮側(cè)索硬化等神經(jīng)退行性疾病直接相關(guān)。
-EAAT5主要在視網(wǎng)膜表達(dá),其氯離子通道特性參與視覺(jué)信號(hào)處理。
-β-內(nèi)酰胺類抗生素(如頭孢曲松)可通過(guò)激活NF-κB通路上調(diào)EAAT2表達(dá),發(fā)揮神經(jīng)保護(hù)作用。
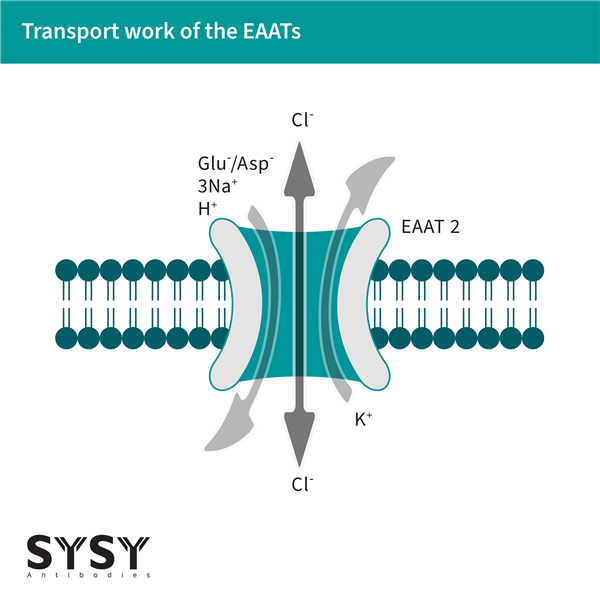
圖1:谷氨酸和陰離子通過(guò)EAATs運(yùn)輸
最近發(fā)表的數(shù)據(jù)表明,EAAT5(谷氨酸/天冬氨酸轉(zhuǎn)運(yùn)蛋白5)是EAAT家族中視網(wǎng)膜特異性的成員,它不僅作為谷氨酸的轉(zhuǎn)運(yùn)蛋白,還在視錐光感受器中作為谷氨酸門(mén)控的Cl?通道發(fā)揮作用。
當(dāng)EAATs將谷氨酸攝取到膠質(zhì)細(xì)胞中時(shí),谷氨酸會(huì)被轉(zhuǎn)化為谷氨酰胺,隨后被運(yùn)回突觸前神經(jīng)元,在那里再轉(zhuǎn)化為谷氨酸,并通過(guò)VGLUTs(囊泡谷氨酸轉(zhuǎn)運(yùn)蛋白)的作用被攝取到突觸小泡中。這一過(guò)程被稱為谷氨酸-谷氨酰胺循環(huán),見(jiàn)圖2。
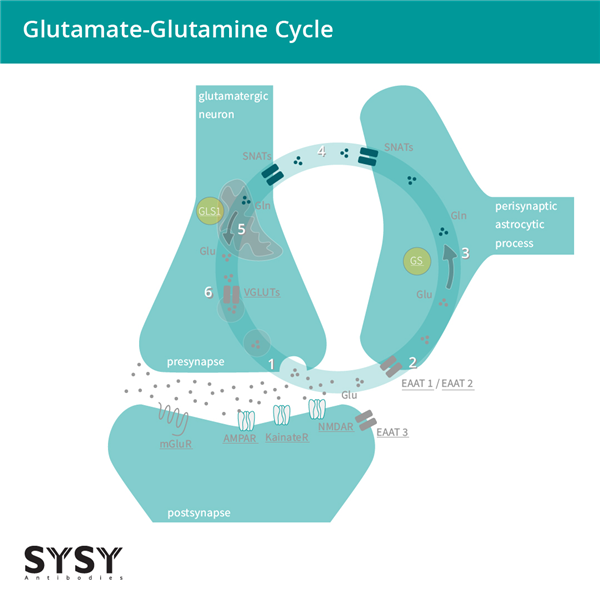
圖2:谷氨酸能神經(jīng)元與星形膠質(zhì)細(xì)胞之間的谷氨酸-谷氨酰胺循環(huán)。
(1) 谷氨酸(Glu)被釋放并結(jié)合到離子型和代謝型受體(AMPA受體/GluA、海人酸受體/GluK、NMDA受體/GluN、mGluRs)。
(2) 谷氨酸主要通過(guò)谷氨酸/天冬氨酸轉(zhuǎn)運(yùn)蛋白(EAAT)1/2被星形膠質(zhì)細(xì)胞攝取,部分通過(guò)EAAT3被神經(jīng)元攝取。
(3) 星形膠質(zhì)細(xì)胞中的谷氨酰胺合成酶將谷氨酸(Glu)轉(zhuǎn)化為谷氨酰胺(Gln)。
(4) 突觸惰性的谷氨酰胺從星形膠質(zhì)細(xì)胞轉(zhuǎn)移到神經(jīng)元。
(5) 谷氨酰胺(Gln)通過(guò)線粒體谷氨酰胺酶1(GLS1)重新轉(zhuǎn)化為谷氨酸(Glu)。
(6) 谷氨酸通過(guò)囊泡谷氨酸轉(zhuǎn)運(yùn)蛋白(VGLUTs)被轉(zhuǎn)運(yùn)到囊泡中,準(zhǔn)備進(jìn)行下一輪的傳遞。
EAATs的分布
EAAT家族目前共有5個(gè)成員,即EAAT1至EAAT5,它們的谷氨酸攝取動(dòng)力學(xué)、氯離子通透性程度以及分布各有不同。
-EAAT1(GLAST):在腦中廣泛分布于星形膠質(zhì)細(xì)胞(包括小腦的Bergmann膠質(zhì)細(xì)胞和視網(wǎng)膜的Müller膠質(zhì)細(xì)胞),是小腦和視網(wǎng)膜中谷氨酸的主要收集者。在小腦皮層等區(qū)域的表達(dá)量較高。
-EAAT2(Glt-1):是除小腦和視網(wǎng)膜外其他腦區(qū)的主要谷氨酸轉(zhuǎn)運(yùn)蛋白,尤其在海馬和皮層中最為突出。在所采樣的所有腦區(qū)中,其表達(dá)量大多處于較高水平,如在視覺(jué)皮層中處于99百分位。
-EAAT3:主要存在于興奮性和抑制性神經(jīng)元的胞體和樹(shù)突中。
-EAAT4和EAAT5:主要在神經(jīng)系統(tǒng)中發(fā)揮作用,且被認(rèn)為主要作為谷氨酸門(mén)控的氯離子通道調(diào)節(jié)神經(jīng)信號(hào)。其中EAAT5是視網(wǎng)膜特異性的成員,主要在視錐光感受器和雙極細(xì)胞等的突觸部位以斑點(diǎn)狀表達(dá),與谷氨酸釋放位點(diǎn)密切相關(guān)。
| 蛋白 | 基因 | 分子量(小鼠) | 氯離子電導(dǎo) | 組織分布 |
| EAT2 | SLC1A2 | ~ 62 kDa | 低 | 主要是星形膠質(zhì)細(xì)胞;介導(dǎo)超過(guò)90%的中樞神經(jīng)系統(tǒng)谷氨酸再攝取 |
| EAT3 | SLC1A1 | ~ 57 kDa | 中等 | 所有神經(jīng)元 - 位于樹(shù)突和軸突末端,腎和胃腸道上皮細(xì)胞(在肺、腎、骨骼肌和小腸中含量低) |
| EAT4 | SLC1A6 | ~ 61 kDa | 高 | 神經(jīng)元(突觸后、樹(shù)突棘)在小腦中 |
| EAT5 | SLC1A7 | ~ 60 kDa | 高 | 中樞神經(jīng)系統(tǒng):主要是視網(wǎng)膜 |
EAAT1:
EAAT1,也被稱為GLAST-1,在中樞神經(jīng)系統(tǒng)(CNS)中廣泛表達(dá),并且在小腦的星形膠質(zhì)細(xì)胞和Bergmann膠質(zhì)細(xì)胞中高表達(dá)。在視網(wǎng)膜中,EAAT1在Müller細(xì)胞中表達(dá)。強(qiáng)大的EAAT表達(dá),特別是EAAT1,是成年神經(jīng)干細(xì)胞(NSC)表型的廣泛使用標(biāo)記。我們提供幾種經(jīng)過(guò)基因敲除(KO)驗(yàn)證的抗體,用于檢測(cè)EAAT1。
| 產(chǎn)品編號(hào) | 產(chǎn)品描述 | 應(yīng)用 | 數(shù)量 |
| 250 103 | EAT1,兔源,多克隆,親和純化(K.O. 細(xì)胞外結(jié)構(gòu)域) | 蛋白質(zhì)印跡(WB) | 50 微克 |
| 250 113 | EAT1,兔源,多克隆,親和純化(K.O. 細(xì)胞質(zhì)結(jié)構(gòu)域) | 蛋白質(zhì)印跡(WB)、免疫沉淀(IP)、細(xì)胞培養(yǎng)(ICC)、免疫組化(IHC) | 50 微克 |
| 250 114 | EAT1,豚鼠源,多克隆,抗血清(細(xì)胞質(zhì)結(jié)構(gòu)域) | 蛋白質(zhì)印跡(WB)、細(xì)胞培養(yǎng)(ICC)、免疫組化(IHC)、免疫組化-過(guò)氧化物酶(IHC-P) | 100 微升 |
| 250 116 | EAT1,雞源,多克隆,IgY組分(細(xì)胞質(zhì)結(jié)構(gòu)域) | 蛋白質(zhì)印跡(WB)、細(xì)胞培養(yǎng)(ICC)、免疫組化(IHC) | 200 微升 |
| 250-11P | EAT1,對(duì)照肽(細(xì)胞質(zhì)結(jié)構(gòu)域) | 100 微克 | |
| 250-1P | EAT1,對(duì)照肽(細(xì)胞外結(jié)構(gòu)域) | 100 微克 |
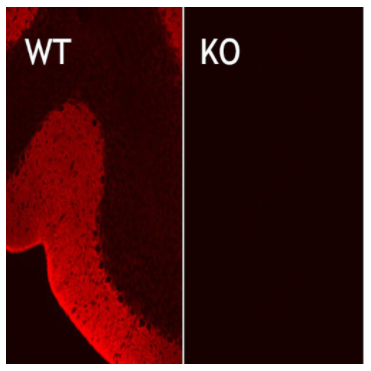
圖3:在野生型(WT)和基因敲除(KO)小鼠小腦中EAAT1的間接免疫染色,使用兔多克隆抗EAAT1(Cat:#250 113,稀釋度1:5000;紅色)。
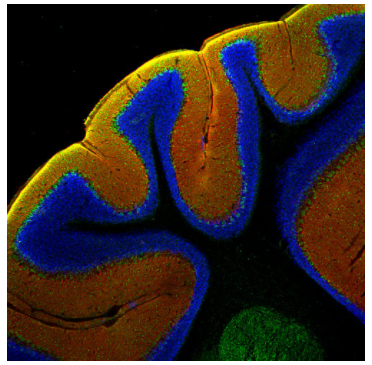
圖4:使用豚鼠多克隆抗EAAT1(Cat:#250 114,稀釋度1:500;紅色)和兔抗parvalbumin(Cat:#195 002,稀釋度1:500;綠色)對(duì)PFA固定的鼠小腦進(jìn)行間接免疫染色。細(xì)胞核通過(guò)DAPI染色顯示(藍(lán)色)。
EAAT2:
EAAT2,也被稱為GLT-1,是大腦中含量最豐富的谷氨酸轉(zhuǎn)運(yùn)蛋白。它主要定位于星形膠質(zhì)細(xì)胞的分支中,并在小腦和海馬中高度表達(dá)。最近的研究發(fā)現(xiàn),在阿爾茨海默病(Alzheimer’s Disease)、多發(fā)性硬化癥(Multiple Sclerosis)和肌萎縮側(cè)索硬化癥(Amyotrophic Lateral Sclerosis, ALS)等多種神經(jīng)退行性疾病中,EAAT2的表達(dá)量有所減少。在此背景下,值得注意的是,γ-分泌酶的活性亞基前體蛋白1(Presenilin 1, PS1)能夠直接與EAAT2相互作用,并影響該轉(zhuǎn)運(yùn)蛋白在細(xì)胞表面的定位。
為了檢測(cè)EAAT2,我們提供了經(jīng)過(guò)基因敲除(KO)驗(yàn)證的優(yōu)質(zhì)多克隆兔和豚鼠抗體。此外,還有一種小鼠單克隆抗體,在Western Blot(WB)、免疫細(xì)胞化學(xué)(ICC)、免疫組織化學(xué)(IHC)和福爾馬林固定石蠟包埋(IHC-P)等應(yīng)用中表現(xiàn)出色。
| 產(chǎn)品編號(hào) | 產(chǎn)品描述 | 應(yīng)用 |
| 250 204 | EAT2,豚鼠源,多克隆,抗血清(K.O. 細(xì)胞外結(jié)構(gòu)域) | 蛋白質(zhì)印跡(WB)、細(xì)胞培養(yǎng)(ICC)、免疫組化(IHC)、免疫組化-過(guò)氧化物酶(IHC-P) |
| 250 211 | EAT2,鼠源,單克隆,純化IgG(細(xì)胞外結(jié)構(gòu)域) | 蛋白質(zhì)印跡(WB)、細(xì)胞培養(yǎng)(ICC)、免疫組化(IHC)、免疫組化-過(guò)氧化物酶(IHC-P) |
| 250-2P | EAT2,對(duì)照肽(細(xì)胞外結(jié)構(gòu)域) |
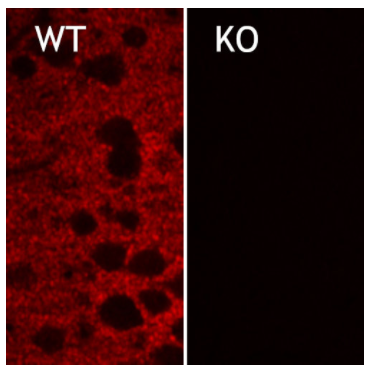
圖5:在雜合子(+/-)和基因敲除(-/-)小鼠的新皮層中EAAT2的間接免疫染色(Cat:#250 203,稀釋度1:2000;紅色)。
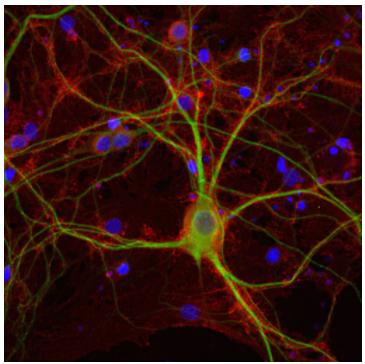
圖6:使用抗EAAT2(Cat:#250 204,稀釋度1:500;紅色)和小鼠抗MAP2(Cat:#188 011,稀釋度1:500;綠色)對(duì)PFA固定的鼠海馬神經(jīng)元進(jìn)行間接免疫染色。細(xì)胞核通過(guò)DAPI染色顯示(藍(lán)色)。
EAAT3:
EAAT3,也被稱為EAAC1或SLC1A1,是大腦中的“神經(jīng)元”谷氨酸轉(zhuǎn)運(yùn)蛋白,主要定位于大腦皮層、海馬、紋狀體和基底神經(jīng)節(jié)的軸突末梢和樹(shù)突。最新的研究結(jié)果表明,EAAT3的表達(dá)與強(qiáng)迫癥(OCD)之間存在聯(lián)系。在中樞神經(jīng)系統(tǒng)之外,EAAT3還存在于肺、小腸、骨骼肌、腎外髓層、髓射線和皮質(zhì)中。
為了檢測(cè)EAAT3,我們提供了兩種出色的多克隆兔抗體。
| 產(chǎn)品編號(hào) | 產(chǎn)品描述 | 應(yīng)用 |
| 250 303 | EAT3,兔源,多克隆,親和純化(目前無(wú)貨) 細(xì)胞質(zhì)結(jié)構(gòu)域 | 細(xì)胞培養(yǎng)(ICC)、免疫組化(IHC) |
| 250 313 | EAT3,兔源,多克隆,親和純化(細(xì)胞質(zhì)結(jié)構(gòu)域) | 蛋白質(zhì)印跡(WB) |
| 250-31P | EAT3,對(duì)照肽(細(xì)胞質(zhì)結(jié)構(gòu)域) |
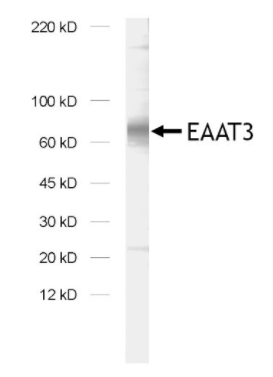
圖7:使用兔多克隆抗EAAT3(Cat:#250 313)對(duì)大鼠腦突觸膜組分(LP1)進(jìn)行免疫印跡分析。
EAAT4:
EAAT4,也被稱為SLC1A6,是一種主要的神經(jīng)元谷氨酸轉(zhuǎn)運(yùn)蛋白。它主要定位于小腦的浦肯野細(xì)胞上,同時(shí)在前腦和中腦的某些亞區(qū)域也有少量表達(dá)。最新的研究表明,EAAT4的表達(dá)呈現(xiàn)出與醛糖酶C(zebrin)相似的矢狀帶狀模式,形成了具有高、低水平EAAT4的分子多樣性浦肯野細(xì)胞微區(qū)域。與EAAT1和EAAT2相比,EAAT4的谷氨酸轉(zhuǎn)運(yùn)能力較低,因此一些研究者推測(cè)EAAT4在生理上主要作為陰離子通道發(fā)揮作用。
我們新推出的兩種針對(duì)EAAT4的兔多克隆抗體表現(xiàn)出色。
| 產(chǎn)品編號(hào) | 產(chǎn)品描述 | 應(yīng)用 |
| 250 403 | EAT4,兔源,多克隆,親和純化 | 蛋白質(zhì)印跡(WB) |
| 250 413 | EAT4,兔源,多克隆,親和純化 | 蛋白質(zhì)印跡(WB)、細(xì)胞培養(yǎng)(ICC)、免疫組化(IHC)、免疫組化-過(guò)氧化物酶(IHC-P) |
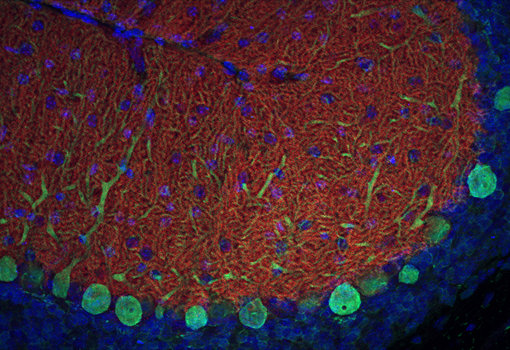
圖8:使用兔抗EAAT4抗體(Cat:#250 413,稀釋度1:500,紅色)和豚鼠抗Calbindin抗體(Cat:#214 318,稀釋度1:500,綠色)對(duì)甲醛固定的鼠小腦切片(矢狀面)進(jìn)行間接免疫染色。細(xì)胞核通過(guò)DAPI染色顯示(藍(lán)色)。
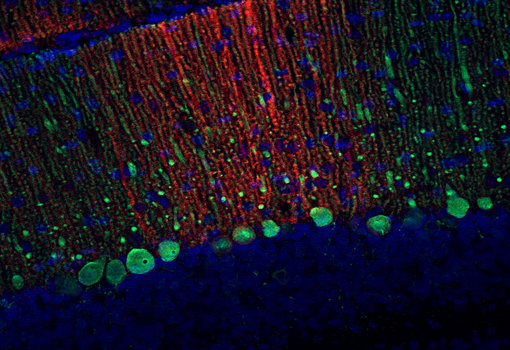
圖9:使用兔抗EAAT4抗體(Cat:#250 413,稀釋度1:500,紅色)和豚鼠抗Calbindin抗體(Cat:#214 318,稀釋度1:500,綠色)對(duì)甲醛固定的鼠小腦切片(冠狀面)進(jìn)行間接免疫染色。細(xì)胞核通過(guò)DAPI染色顯示(藍(lán)色)。
EAAT5:
EAAT5蛋白,有時(shí)也被稱為AAAT,主要存在于脊椎動(dòng)物的視網(wǎng)膜中,它與EAAT2一起將谷氨酸轉(zhuǎn)運(yùn)到視桿細(xì)胞、視錐細(xì)胞和視桿雙極細(xì)胞中。EAAT5轉(zhuǎn)運(yùn)蛋白聚集在視桿和視錐細(xì)胞突觸前活性區(qū)的突觸帶下方,正好位于谷氨酸釋放的區(qū)域,能夠高效地捕獲剛剛釋放的谷氨酸。抑制EAAT5的研究表明,這種蛋白對(duì)于視網(wǎng)膜中甘氨酸能無(wú)長(zhǎng)突細(xì)胞(AII無(wú)長(zhǎng)突細(xì)胞)的時(shí)間信號(hào)分辨率非常重要。在中樞神經(jīng)系統(tǒng)之外,EAAT5還在肝臟、腎臟、腸道、心臟、肺和肌肉中表達(dá)。
我們提供了一種經(jīng)過(guò)基因敲除(KO)驗(yàn)證的豚鼠多克隆抗EAAT5抗體。基因敲除驗(yàn)證是一種確保抗體特異性的方法,通過(guò)CRISPR-Cas9基因組編輯技術(shù),驗(yàn)證抗體是否能夠特異性地識(shí)別其靶標(biāo)蛋白。這種驗(yàn)證方法可以提高抗體在研究中的可靠性和可重復(fù)性。
| 產(chǎn)品編號(hào) | 產(chǎn)品描述 | 應(yīng)用 |
| 250 504 | EAT5,豚鼠源,多克隆,抗血清 | 免疫組化(IHC)、免疫組化-過(guò)氧化物酶(IHC-P) |
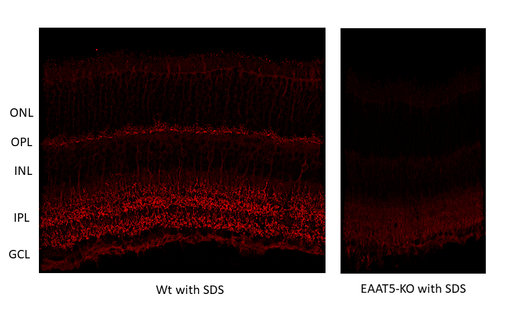
圖10:使用兔抗EAAT5抗體(Cat:#250 504,稀釋度1:2000;紅色)對(duì)野生型(WT)和基因敲除(KO)小鼠視網(wǎng)膜進(jìn)行間接免疫染色。組織經(jīng)4%甲醛固定,,使用1% SDS進(jìn)行抗原修復(fù)。
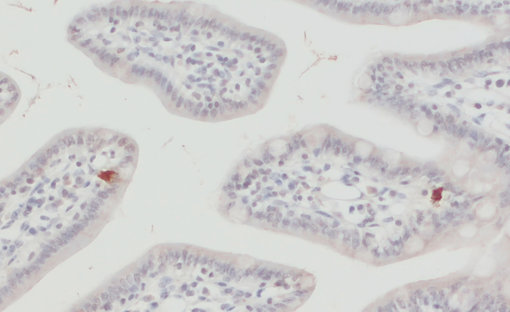
圖11:使用豚鼠抗EAAT5抗體(Cat:#250 504,稀釋度1:1000,DAB;棕色)對(duì)甲醛固定的石蠟包埋(FFPE)小鼠回腸切片進(jìn)行間接免疫染色。細(xì)胞核通過(guò)蘇木精染色顯示(藍(lán)色)。
產(chǎn)品訂購(gòu):sales@amyjet.com
郵政編碼:430070
公司地址:武漢市洪山區(qū)光谷大道35號(hào)
光谷總部國(guó)際二期時(shí)代1棟13樓
提示:本公司所有產(chǎn)品僅供科研使用,不用于臨床診斷。
版權(quán)所有:艾美捷科技有限公司 鄂ICP備10204150號(hào)-1 鄂公網(wǎng)安備:42018502004523號(hào)
第二類醫(yī)療器械經(jīng)營(yíng)備案憑證:鄂漢藥監(jiān)械經(jīng)營(yíng)備20234324號(hào)
友情鏈接:亞科因生物每日生物評(píng)論

微信掃碼在線客服