PRODUCT CENTER
特色產(chǎn)品當(dāng)前位置:首頁(yè) > 特色產(chǎn)品
神經(jīng)肽是大腦中最大、最多樣化的信號(hào)分子類別。它們可以直接作為神經(jīng)遞質(zhì),作為其他遞質(zhì)傳遞的調(diào)節(jié)劑,作為緊密細(xì)胞環(huán)境中的自分泌或旁分泌調(diào)節(jié)劑,以及作為長(zhǎng)期激素。神經(jīng)肽由神經(jīng)元合成和/或使用。它們最初作為大的不活躍前體蛋白(稱為前體肽)合成。前體肽包含一個(gè)N端信號(hào)肽,這是新合成基因產(chǎn)物進(jìn)入內(nèi)質(zhì)網(wǎng)腔所必需的。信號(hào)序列在通過內(nèi)質(zhì)網(wǎng)膜時(shí)被切除,形成內(nèi)質(zhì)網(wǎng)-高爾基體中的前肽,進(jìn)一步排序進(jìn)入調(diào)節(jié)分泌途徑。為了釋放生物活性神經(jīng)肽,前肽進(jìn)一步被內(nèi)肽酶和外肽酶切割,并進(jìn)行翻譯后修飾,例如糖基化、磷酸化、硫酸化、乙酰化、添加寡糖和N端吡咯谷氨酸形成。最常見的修飾是C端酰胺化,大約一半的已知生物活性肽是α-酰胺化的(圖1)。
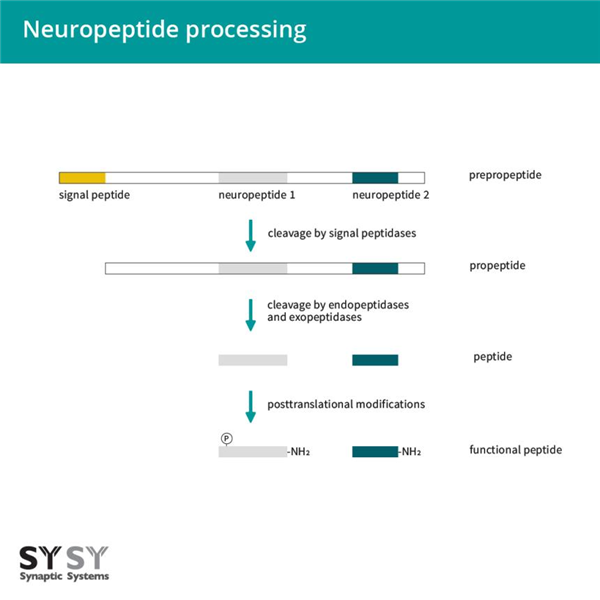
圖1:神經(jīng)肽加工。神經(jīng)肽作為大的不活躍前體蛋白的一部分合成。幾個(gè)蛋白水解切割步驟和翻譯后修飾導(dǎo)致生物活性肽的產(chǎn)生。
內(nèi)蛋白水解切割和翻譯后修飾發(fā)生在高爾基體網(wǎng)絡(luò)和肽包裝的致密核心囊泡中。致密核心囊泡在整個(gè)神經(jīng)元中運(yùn)輸,并可以在突觸間隙、細(xì)胞體和沿軸突釋放肽(圖2)。
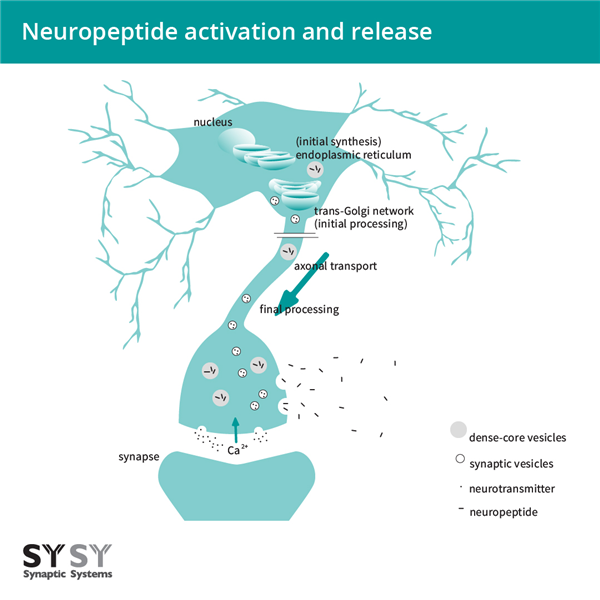
圖2:神經(jīng)肽激活和釋放。神經(jīng)肽最初在內(nèi)質(zhì)網(wǎng)中合成,在高爾基體網(wǎng)絡(luò)和致密核心囊泡中被切割和翻譯后修飾,肽被包裝在其中。神經(jīng)肽在致密核心囊泡中沿軸突運(yùn)輸,鈣離子流入時(shí)釋放,并通過擴(kuò)散分布。
神經(jīng)肽的釋放、擴(kuò)散和失活
神經(jīng)肽通常與同一神經(jīng)元中的其他神經(jīng)肽和神經(jīng)遞質(zhì)共同釋放,產(chǎn)生多種效果。在突觸中,致密核心囊泡與含有經(jīng)典神經(jīng)遞質(zhì)如谷氨酸的突觸囊泡共定位。盡管致密核心囊泡和突觸囊泡通常共同釋放,但它們使用不同的機(jī)制。像神經(jīng)遞質(zhì)一樣,神經(jīng)肽通過鈣依賴性胞吐作用釋放,以響應(yīng)去極化或其他信號(hào)。然而,來自致密核心囊泡的神經(jīng)肽在比突觸囊泡中的神經(jīng)遞質(zhì)更低的細(xì)胞質(zhì)[Ca2+]濃度下釋放。傳統(tǒng)神經(jīng)遞質(zhì)的釋放被認(rèn)為發(fā)生在非常接近Ca2+進(jìn)入部位的地方,而神經(jīng)肽通常在遠(yuǎn)離Ca2+進(jìn)入部位的地方釋放。因此,致密核心囊泡相對(duì)于Ca2+流入部位的位置可能決定了分泌所需的Ca2+量。
與經(jīng)典神經(jīng)遞質(zhì)相比,神經(jīng)肽從其釋放部位擴(kuò)散,因此可以在相對(duì)較大的范圍內(nèi)(納米到毫米)起作用(圖2)。這種擴(kuò)散驅(qū)動(dòng)的分布被稱為體積傳遞或擴(kuò)散。
由于沒有肽的再攝取機(jī)制,它們只能緩慢地從細(xì)胞外空間中移除。相比之下,經(jīng)典神經(jīng)遞質(zhì)通過專門的轉(zhuǎn)運(yùn)蛋白迅速?gòu)耐挥|間隙中移除。體積傳遞和缺乏再攝取的結(jié)合導(dǎo)致神經(jīng)肽的相對(duì)持久效果。與神經(jīng)遞質(zhì)相比,神經(jīng)肽壽命長(zhǎng),但其效果會(huì)終止。失活通過細(xì)胞外蛋白酶發(fā)生,這在某些情況下甚至可以通過切割現(xiàn)有神經(jīng)肽生成新的生物活性肽。
所有神經(jīng)肽都通過細(xì)胞表面受體作為信號(hào)轉(zhuǎn)導(dǎo)體。幾乎所有神經(jīng)肽都作用于G蛋白偶聯(lián)受體,觸發(fā)第二信使級(jí)聯(lián)反應(yīng)以調(diào)節(jié)細(xì)胞活動(dòng)。像肽配體一樣,受體不僅在神經(jīng)系統(tǒng)中廣泛分布,而且在許多其他組織中也廣泛分布。神經(jīng)肽受體與神經(jīng)遞質(zhì)受體相比具有相對(duì)較高的配體親和力(納摩爾Kds)(微摩爾Kds)。這樣,少量擴(kuò)散的肽仍然可以激活受體。這一事實(shí)及其長(zhǎng)壽命使神經(jīng)肽能夠在相對(duì)較低的濃度下在相對(duì)較遠(yuǎn)的距離上活躍。
差異表達(dá)和加工多樣性
由于可變剪接、串聯(lián)組織或細(xì)胞特異性分化的翻譯后加工前肽,單個(gè)神經(jīng)肽基因通常表現(xiàn)出多種表型。可變剪接是在顯示降鈣素基因產(chǎn)生編碼降鈣素肽或降鈣素基因相關(guān)肽(CGRPs)的mRNA時(shí)發(fā)現(xiàn)的。對(duì)于一些神經(jīng)肽,前肽的差異加工產(chǎn)生不同長(zhǎng)度的成熟肽,這些肽具有相同的受體結(jié)合表位(圖3A)。盡管同一前體的不同產(chǎn)物結(jié)合到同一受體,但它們從循環(huán)中的不同清除率影響它們的效果。因此,proCCK主要加工成CCK-33、CCK-12或CCK-8,或者prosomatostatin加工成somatostatin-28或somatostatin-14是相關(guān)的。
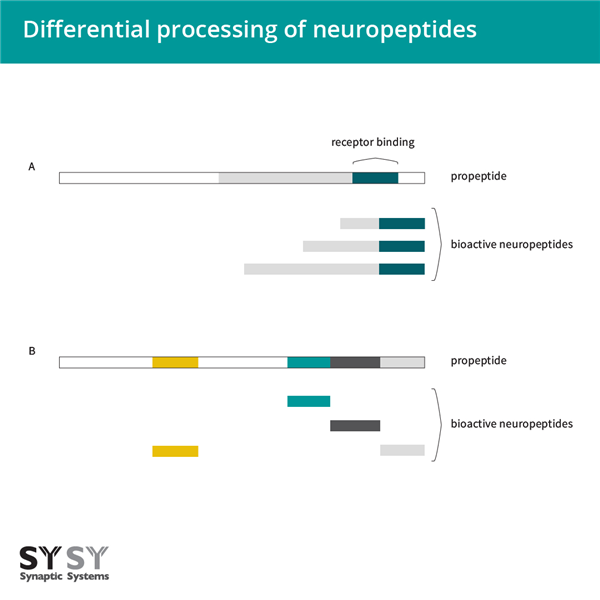
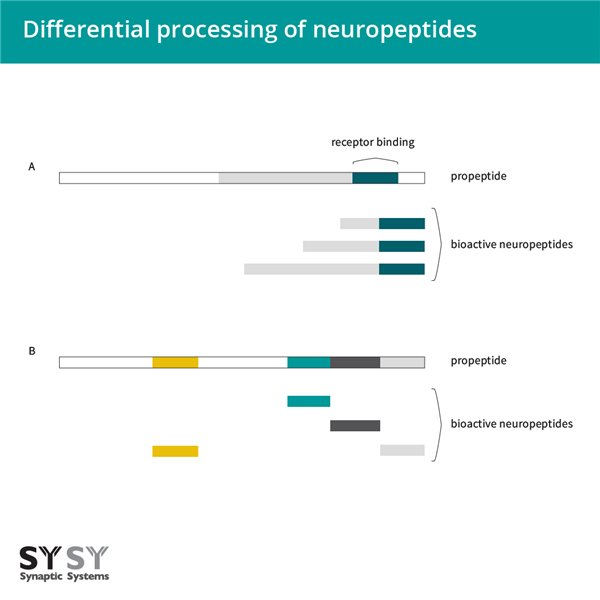
圖3:神經(jīng)肽的差異加工A:對(duì)于一些神經(jīng)肽,前肽的差異加工導(dǎo)致成熟肽的不同長(zhǎng)度,但仍然共享相同的受體結(jié)合表位;B:在某些情況下,前肽包含不同的神經(jīng)肽,可以在不同組織中差異加工。
基因表達(dá)不同生物活性肽的另一種方式是當(dāng)基因本身編碼一個(gè)包含不同神經(jīng)肽的前肽(圖3B),例如阿片肽基因和一些速激肽基因。這些同一前肽中的各種神經(jīng)肽可以在不同組織中差異加工。神經(jīng)肽在各種目標(biāo)組織中起作用。它們的作用可以是局部的或遠(yuǎn)距離的。因此,幾乎所有的身體功能都可以被調(diào)節(jié)。許多具有相似結(jié)構(gòu)的神經(jīng)肽具有非常不同的功能。例如,加壓素和催產(chǎn)素是兩種下丘腦肽,每種由九個(gè)氨基酸組成(見圖4A和4B的腦切片IHC染色)。這兩種肽在這九個(gè)殘基中有七個(gè)是相同的,被認(rèn)為是進(jìn)化早期基因重復(fù)的結(jié)果。兩種肽的作用是不同的:催產(chǎn)素引起乳汁釋放和子宮收縮,而加壓素引起腎臟和血管收縮。
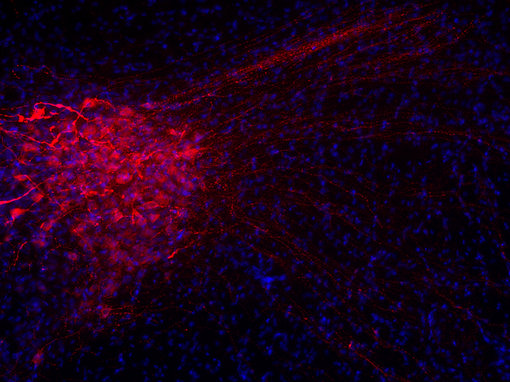
圖4A:使用豚鼠抗加壓素抗體(403 004,稀釋1:500,紅色)對(duì)PFA固定的大鼠下丘腦切片進(jìn)行間接免疫染色。細(xì)胞核通過DAPI染色(藍(lán)色)可視化。
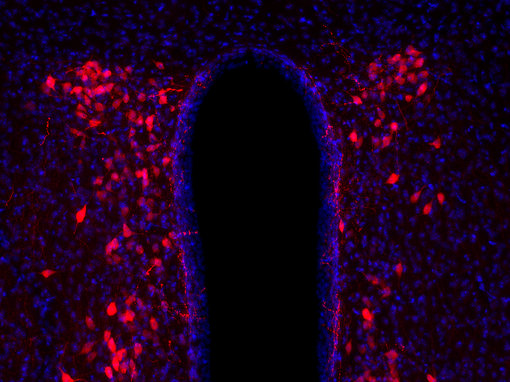
圖4B:使用豚鼠抗催產(chǎn)素抗體(408 004,稀釋1:500,紅色)對(duì)PFA固定的小鼠下丘腦切片進(jìn)行間接免疫染色。細(xì)胞核通過DAPI染色(藍(lán)色)可視化。
神經(jīng)肽在疼痛傳遞中一直受到關(guān)注。對(duì)轉(zhuǎn)基因小鼠的研究表明,缺乏物質(zhì)P或其受體的小鼠對(duì)中度或重度疼痛沒有反應(yīng)。另一種肽,降鈣素基因相關(guān)肽(CGRP),在偏頭痛的病理生理中起重要作用(見圖5A和5B的脊髓切片染色)。
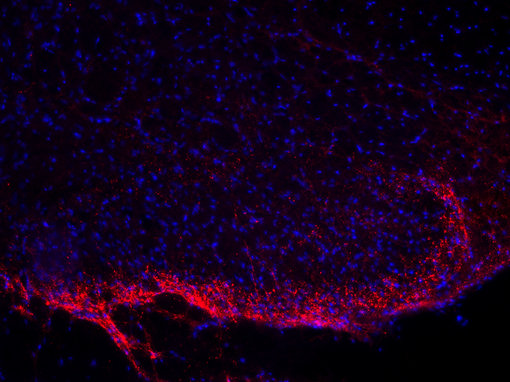
圖5A:使用豚鼠抗物質(zhì)P抗體(459 005,稀釋1:500,紅色)對(duì)PFA固定的大鼠脊髓切片進(jìn)行間接免疫染色。細(xì)胞核通過DAPI染色(藍(lán)色)可視化。
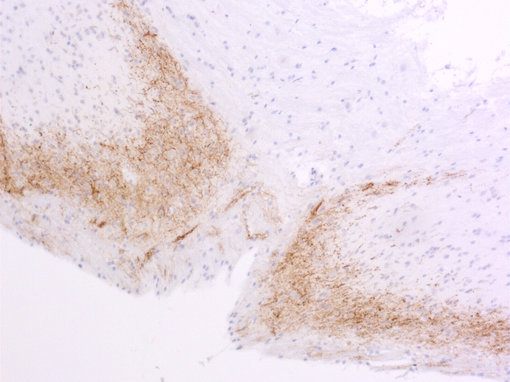
圖5B:使用豚鼠抗CGRP抗體(414 004,稀釋1:1000,DAB)對(duì)PFA固定的石蠟包埋大鼠脊髓切片進(jìn)行間接免疫染色。細(xì)胞核用蘇木精(藍(lán)色)復(fù)染。
中樞神經(jīng)系統(tǒng)對(duì)食物攝入的控制是另一個(gè)正在進(jìn)行的研究課題。神經(jīng)肽Y刺激碳水化合物攝入,而加拉寧刺激脂肪攝入(見圖6A和6B的腦切片IHC染色)。Agouti相關(guān)肽和食欲素也有刺激作用。其他神經(jīng)肽,如黑素皮質(zhì)素和可卡因和安非他明調(diào)節(jié)轉(zhuǎn)錄本,抑制食物攝入。
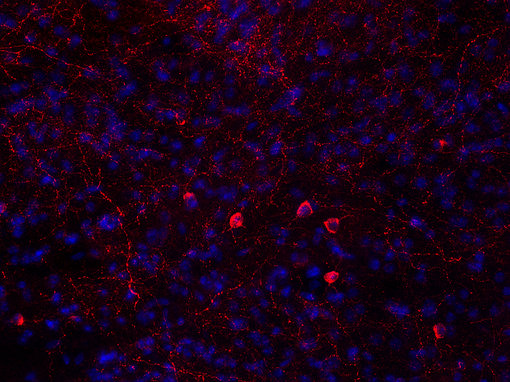
圖6A:使用雞抗神經(jīng)肽Y抗體(394 006,稀釋1:500,紅色)對(duì)PFA固定的小鼠紋狀體切片進(jìn)行間接免疫染色。細(xì)胞核通過DAPI染色(藍(lán)色)可視化。在染色前應(yīng)用抗原修復(fù)(10mM Tris,1mM EDTA,pH 9.0,60°C過夜)。
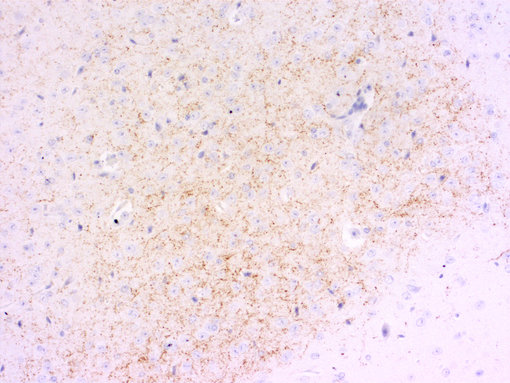
圖6B:使用豚鼠抗加拉寧抗體(446 004,稀釋1:1000,DAB)對(duì)PFA固定的石蠟包埋小鼠下丘腦切片進(jìn)行間接免疫染色。細(xì)胞核用蘇木精(藍(lán)色)復(fù)染。
總之,神經(jīng)肽的各種功能與這類信號(hào)分子本身一樣多樣化。下表列出了一些神經(jīng)肽及其功能:
神經(jīng)肽 | 功能 |
ACTH | 刺激皮質(zhì)醇的產(chǎn)生和釋放 |
AGRP | 刺激食欲和調(diào)節(jié)代謝及能量消耗 |
CART | 調(diào)節(jié)進(jìn)食、獎(jiǎng)勵(lì)和壓力,并作為精神興奮劑 |
CCK-8 | 參與消化、食物攝入、焦慮和恐懼 |
CGRP | 作為血管擴(kuò)張劑和痛覺傳導(dǎo) |
CRF | 刺激ACTH產(chǎn)生,決定妊娠期長(zhǎng)度和分娩時(shí)間 |
Galanin | 參與調(diào)節(jié)進(jìn)食、內(nèi)臟穩(wěn)態(tài)、痛覺、喚醒/睡眠和認(rèn)知 |
神經(jīng)肽S | 參與調(diào)節(jié)喚醒、焦慮和恐懼、食物攝入、學(xué)習(xí)和記憶 |
神經(jīng)肽Y | 參與食物攝入、能量?jī)?chǔ)存、減輕壓力、焦慮和疼痛感知、血壓調(diào)節(jié) |
神經(jīng)緊張素 | 調(diào)節(jié)多巴胺通路、疼痛、體溫、食欲、脂肪代謝和學(xué)習(xí)和記憶 |
Orexin | 調(diào)節(jié)進(jìn)食、睡眠、喚醒和能量穩(wěn)態(tài) |
Oxytocin | 刺激分娩和哺乳期間的平滑肌收縮,具有社會(huì)聯(lián)系和繁殖功能 |
生長(zhǎng)抑素 | 內(nèi)分泌激素分泌的負(fù)調(diào)節(jié)劑 |
物質(zhì)P | 腸道平滑肌收縮、血管舒張、中樞疼痛處理、神經(jīng)源性炎癥、焦慮和壓力 |
血管加壓素 | 調(diào)節(jié)水穩(wěn)態(tài)、血壓和社會(huì)行為 |
VIP | 刺激心臟收縮、血管舒張、調(diào)節(jié)血壓和放松氣管、胃和膽囊的平滑肌 |
疾病和藥物開發(fā)
大量的神經(jīng)肽和神經(jīng)肽受體為藥物靶點(diǎn)發(fā)現(xiàn)提供了許多機(jī)會(huì)。食欲素神經(jīng)元僅位于下丘腦外側(cè),并廣泛分布于大腦(見圖7A和7B的IHC圖片)。
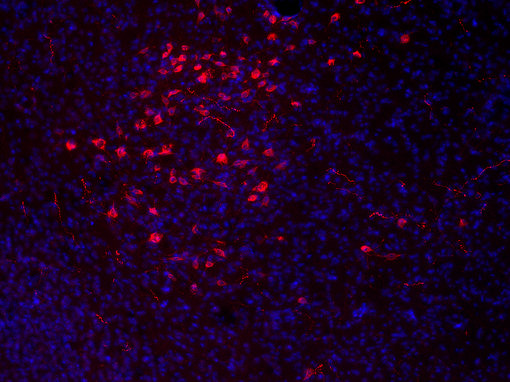
圖7A:使用豚鼠抗食欲素A抗體(389 004,稀釋1:500,紅色)對(duì)PFA固定的小鼠下丘腦切片進(jìn)行間接免疫染色。細(xì)胞核通過DAPI染色(藍(lán)色)可視化。
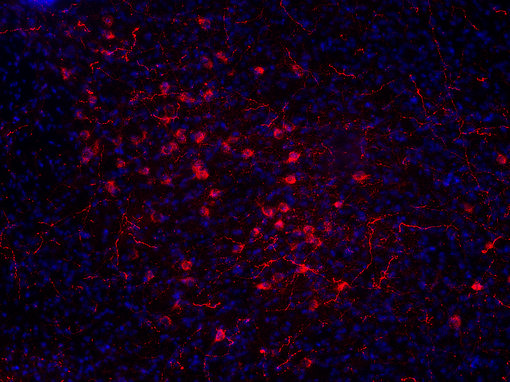
圖7B:使用豚鼠抗食欲素A/B抗體(389 104,稀釋1:500,紅色)對(duì)PFA固定的小鼠下丘腦切片進(jìn)行間接免疫染色。細(xì)胞核通過DAPI染色(藍(lán)色)可視化。
在發(fā)現(xiàn)物質(zhì)P七十年后,第一種肽藥物,一種物質(zhì)P拮抗劑,被臨床測(cè)試用于治療抑郁癥。肽研究進(jìn)展緩慢部分是由于合成選擇性和強(qiáng)效的血腦屏障穿透激動(dòng)劑或拮抗劑的困難。最近,針對(duì)CGRP及其受體的單克隆抗體被引入作為偏頭痛的新治療方法。它們目前是通過阻斷CGRP信號(hào)在偏頭痛預(yù)防中的最新技術(shù)。
針對(duì)神經(jīng)肽的抗體
一般來說,我們旨在開發(fā)針對(duì)活性肽的抗體,并使用加工肽或切割活性肽的末端部分,包括已知修飾,用于免疫。我們的抗體在免疫組織化學(xué)(IHC和IHC-P,如圖8A和8B所示)或免疫細(xì)胞化學(xué)(ICC,圖8C和8D)中表現(xiàn)出色!
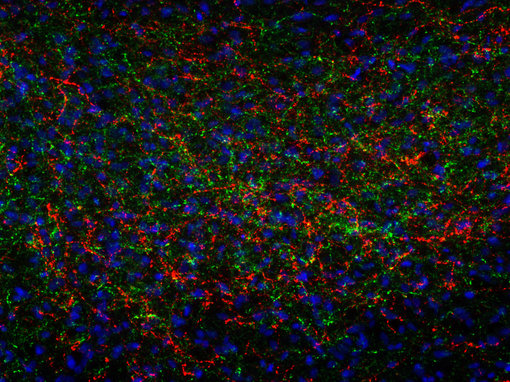
圖8A:使用豚鼠抗ACTH抗體(452 005,稀釋1:500,紅色)和雞抗神經(jīng)肽Y抗體(394 006,稀釋1:500,綠色)對(duì)PFA固定的大鼠下丘腦切片進(jìn)行間接免疫染色。細(xì)胞核通過DAPI染色(藍(lán)色)可視化。
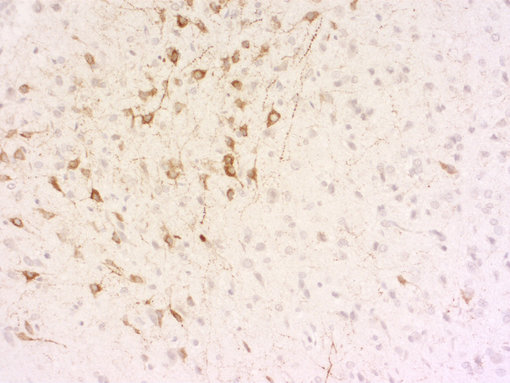
圖8B:使用豚鼠抗生長(zhǎng)抑素-28抗體(366 004,稀釋1:500,DAB)對(duì)PFA固定的石蠟包埋大鼠下丘腦切片進(jìn)行間接免疫染色。細(xì)胞核通過蘇木精染色(藍(lán)色)可視化。
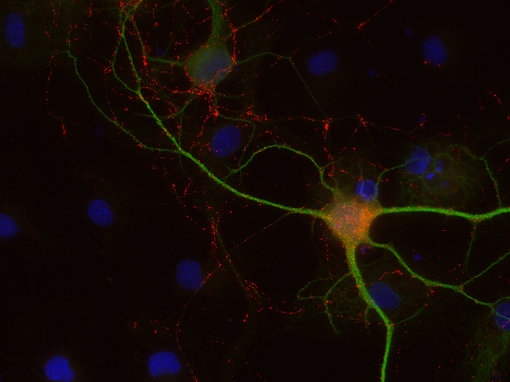
圖8C:使用豚鼠抗CCK-8抗體(438 004,稀釋1:500,紅色)和兔抗MAP 2抗體(188 002,稀釋1:1000,綠色)對(duì)PFA固定的大鼠海馬神經(jīng)元進(jìn)行間接免疫染色。細(xì)胞核通過DAPI染色(藍(lán)色)可視化。
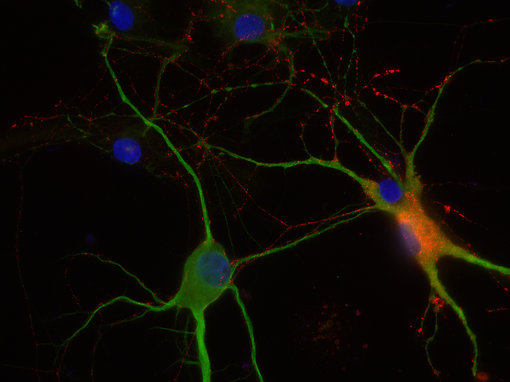
圖8D:使用豚鼠抗VIP抗體(443 005,稀釋1:100,紅色)和兔抗MAP 2抗體(188 002,稀釋1:1000,綠色)對(duì)PFA固定的大鼠海馬神經(jīng)元進(jìn)行間接免疫染色。細(xì)胞核通過DAPI染色(藍(lán)色)可視化。
產(chǎn)品 神經(jīng)肽和肽激素
Cat. No. | Product Description | Application | Quantity |
452 005 | ACTH, Guinea pig, polyclonal, affinity purified | IHC IHC-P | 50 ug |
438 004 | CCK-8, Guinea pig, polyclonal, antiserum | ICC IHC IHC-P | 100 ul |
414 004 | CGRP, Guinea pig, polyclonal, antiserum | IHC IHC-P iDISCO Clarity | 100 ul |
259 002 | Chromogranin A, rabbit, polyclonal, antiserum | WB | 200 ul |
259 003 | Chromogranin A, rabbit, polyclonal, affinity purified (K.O.) | WB ICC IHC IHC-P | 50 ug |
259-0P | Chromogranin A, control protein | - | 100 ug |
259 103 | Chromogranin B, rabbit, polyclonal, affinity purified (K.O.) | WB ICC IHC IHC-P | 50 ug |
259-1P | Chromogranin B, control protein | - | 100 ug |
446 004 | Galanin, Guinea pig, polyclonal, antiserum | ICC IHC IHC-P | 100 ul |
468 003 | Ghrelin, rabbit, polyclonal, affinity purified | IHC IHC-P | 50 ug |
460 003 | GIP, rabbit, polyclonal, affinity purified discontinued, replacement: 514 003 | IHC IHC-P | 50 ug |
514 003 | GIP, rabbit, polyclonal, affinity purified | IHC IHC-P | 50 ug |
471 005 | GLP-1, Guinea pig, polyclonal, affinity purified | Dot blot IHC IHC-P | 50 ug |
471 203 | GLP-2, rabbit, polyclonal, affinity purified | Dot blot IHC IHC-P | 50 ug |
434 005 | Neuropeptide S, Guinea pig, polyclonal, affinity purified | IHC IHC-P | 50 ug |
產(chǎn)品訂購(gòu):sales@amyjet.com
郵政編碼:430070
公司地址:武漢市洪山區(qū)光谷大道35號(hào)
光谷總部國(guó)際二期時(shí)代1棟13樓
提示:本公司所有產(chǎn)品僅供科研使用,不用于臨床診斷。
版權(quán)所有:艾美捷科技有限公司 鄂ICP備10204150號(hào)-1 鄂公網(wǎng)安備:42018502004523號(hào)
第二類醫(yī)療器械經(jīng)營(yíng)備案憑證:鄂漢藥監(jiān)械經(jīng)營(yíng)備20234324號(hào)
友情鏈接:亞科因生物每日生物評(píng)論

微信掃碼在線客服