TECHNICAL COLUMN
學(xué)習(xí)資源當(dāng)前位置:首頁 > 學(xué)習(xí)資源
Argonaute蛋白在進(jìn)化過程中演變出了各種亞科蛋白。這些亞科蛋白可以識別各種不同類型的小RNA分子,從而在各種小RNA沉默途徑中發(fā)揮作用。在經(jīng)典的由siRNA分子介導(dǎo)的RNAi途徑中,Argonaute蛋白可以用內(nèi)切核酸酶活性來沉默mRNA靶分子,這種過程被稱作切割。在生殖細(xì)胞中,面對各種外來的遺傳物質(zhì),Argonaute亞科蛋白PIWI蛋白在piRNA介導(dǎo)的RNA沉默途徑中,利用的也是切割機(jī)制。
在真核生物Ago (eAgo)蛋白,Argonaute (Ago)蛋白可以結(jié)合非編碼的小RNA,參與降解或者抑制互補(bǔ)配對的靶向RNA,從而影響基因表達(dá),稱之為RNA干擾 (RNAi)【2】。揭示RNA干擾機(jī)理的兩位美國科學(xué)家獲得了2006年的諾貝爾生理醫(yī)學(xué)獎。所有的eAgo都具有非常保守的結(jié)構(gòu)功能域,主要由N-結(jié)構(gòu)域(結(jié)合與分離靶向鏈),PAZ結(jié)構(gòu)域(結(jié)合引導(dǎo)鏈的3’末端),MID結(jié)構(gòu)域(結(jié)合引導(dǎo)鏈的5’末端)和PIWI結(jié)構(gòu)域(核酸酶催化中心所在)組成。
細(xì)菌Ago(pAgo)蛋白主要分為長pAgo和短pAgo蛋白。其中,研究較多的長pAgo結(jié)構(gòu)域和eAgo結(jié)構(gòu)域非常相似,主要使用RNA或者DNA作為靶向序列參與清除入侵的核酸(噬菌體或質(zhì)粒)或解除基因組連接。而占已知pAgo大多數(shù)的短pAgo的研究較少。短pAgo只編碼MID結(jié)構(gòu)域和PIWI結(jié)構(gòu)域,并且其PIWI結(jié)構(gòu)域沒有核酸酶催化中心,不能降解結(jié)合的核酸。但是短pAgo通常與一個伴侶蛋白形成異源復(fù)合物發(fā)揮功能。長久以來,細(xì)菌中的短pAgo(約占Ago家族58%)如何發(fā)揮作用以及怎么發(fā)揮作用是領(lǐng)域內(nèi)亟待解決的重大問題【3】。
這一領(lǐng)域的第一個突破性發(fā)現(xiàn)來自荷蘭瓦赫寧根大學(xué)的Daan C. Swarts團(tuán)隊(duì)。2022年,他們研究了一種短pAgo與伴侶蛋白(TIR-APAZ)組成的異源二聚系統(tǒng)MapSPARTA。當(dāng)細(xì)菌遭受外源質(zhì)粒侵襲后,MapSPARTA被激活,在單鏈RNA引導(dǎo)下結(jié)合靶向單鏈DNA后,MapSPARTA可以形成四聚體,從而激活伴侶蛋白中的TIR結(jié)構(gòu)域的NAD(P)+酶活性,進(jìn)而消耗細(xì)菌體內(nèi)的NAD(P)+。因?yàn)镹ADP是維持細(xì)菌生存必需的小分子,NADP的耗竭將會導(dǎo)致細(xì)菌死亡。這樣,通過犧牲侵染細(xì)菌個體達(dá)到保護(hù)群體的效果【4】。Swarts團(tuán)隊(duì)的發(fā)現(xiàn)未能揭示MapSPARTA的結(jié)構(gòu)和活化機(jī)理。
2023年7月26日,來自俄亥俄州立大學(xué)的傅天民團(tuán)隊(duì),在Nature以加速出版形式發(fā)表論文Oligomerization-mediated activation of a short prokaryotic Argonaute,他們利用冷凍電鏡和生物化學(xué)手段,完整揭示了的MapSPARTA活化機(jī)理。

在文中,作者捕捉到一系列高分辨率的MapSPARTA構(gòu)象,包括未激活狀態(tài)下的MapSPARTA單體結(jié)構(gòu),結(jié)合引導(dǎo)RNA鏈和靶向DNA鏈后的單體結(jié)構(gòu),MapSPARTA二聚化的中間態(tài)結(jié)構(gòu),四聚化的活化態(tài)的MapSPARTA,以及四聚體與NAD+的復(fù)合物,完整呈現(xiàn)了MapSPARTA獨(dú)特的層級激活分子機(jī)制。鑒于Ago蛋白在眾多生物體中的廣泛存在,研究清楚MapSPARTA的作用機(jī)制,不僅有助于理解短pAgo的生物學(xué)功能和分子作用機(jī)制,也為改造pAgo奠定了基礎(chǔ)。具體研究內(nèi)容如下:
1)解析了分辨率為3.1 ?的未激活狀態(tài)下的MapSPARTA單體結(jié)構(gòu),闡明了單體的基本結(jié)構(gòu)組成,以及各個結(jié)構(gòu)域的相互作用。
2)與其他Ago相比,MapSPARTA獨(dú)特的APAZ結(jié)構(gòu)域有一個C末端基序,這個C末端基序占據(jù)核酸結(jié)合口袋,起到抑制了核酸結(jié)合的功能。
3)闡明了MapSPARTA識別RNA引導(dǎo)鏈和DNA靶向鏈的分子基礎(chǔ)。與其他Ago相比,短的pAgoMID結(jié)構(gòu)域中特別存在的兩個Loop,導(dǎo)致RNA引導(dǎo)鏈和DNA靶向鏈從第三位才開始互補(bǔ)配對。
4)解析了分辨率為2.7 ?的活化狀態(tài)下的四聚體MapSPARTA結(jié)構(gòu),結(jié)構(gòu)像一個美麗的蝴蝶。四聚體中存在兩類相互作用:一類通過兩個MID-MID結(jié)構(gòu)域介導(dǎo);另一類則是通過4個TIR結(jié)構(gòu)域形成聚集體來實(shí)現(xiàn)的。四個MapSPARTA形成一個不對稱的四聚體。四個SPARTA分子有兩種不同的狀態(tài),兩種狀態(tài)下的TIR結(jié)構(gòu)域有大約180度的翻轉(zhuǎn)。
5)TIR四聚體中有兩種相互作用界面:二聚化作用界面和四聚化作用界面。TIR結(jié)構(gòu)域可以催化NAD降解,研究表明NAD+分子結(jié)合在TIR結(jié)構(gòu)域兩個二聚化作用界面上。進(jìn)一步生化分析發(fā)現(xiàn)四聚體對于TIR的催化是必要的。
6)本文捕捉到了MapSPARTA未激活結(jié)構(gòu),結(jié)合核酸的單體結(jié)構(gòu),結(jié)合核酸的二聚體結(jié)構(gòu), 以及活化的四聚體結(jié)構(gòu),這一系列結(jié)構(gòu)揭示了MapSPARTA的層級激活機(jī)理。未激活狀態(tài)下的MapSPARTA單體結(jié)合RNA引導(dǎo)鏈/DNA靶向鏈后,MID結(jié)構(gòu)域發(fā)生構(gòu)象變化。接著,兩個結(jié)合互補(bǔ)核酸鏈的MapSPARTA單體相互靠近后,由于MID形狀和電荷的互補(bǔ),MID結(jié)構(gòu)域和另一個MID結(jié)構(gòu)域發(fā)生相互作用,形成一個對稱的二聚體。在對稱二聚體中,APAZ連接的TIR結(jié)構(gòu)域由于電荷和形狀的互斥反應(yīng),其中一個TIR結(jié)構(gòu)域發(fā)生近約180度的翻轉(zhuǎn),形成不對稱二聚體。最后兩個不對稱二聚化的MapSPARTA再通過TIR相互作用變?yōu)樗木刍せ顮顟B(tài)。
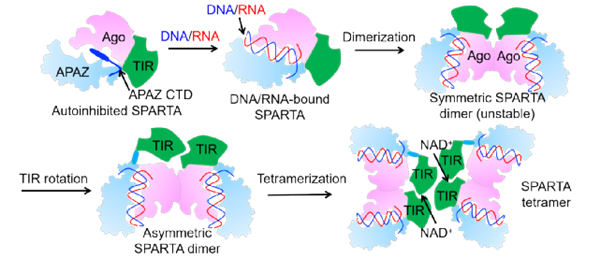
圖1. MapSPARTA的圖示激活過程
綜上,本研究首次從分子水平上描述了短pAgo家族MapSPARTA的激活機(jī)制,為理解其他短Ago活化機(jī)理奠定了基礎(chǔ),也為進(jìn)一步改造MapSPARTA成為核酸檢測或治療工具提供了理論支持。
原文鏈接:
https://www.nature.com/articles/s41586-023-06456-z
參考文獻(xiàn):
1.Swarts, D. C. et al. The evolutionary journey of Argonaute proteins. Nat Struct Mol Biol 21, 743-753, doi:10.1038/nsmb.2879 (2014).
2.Peters, L. & Meister, G. Argonaute proteins: mediators of RNA silencing. Mol Cell 26, 611-623, doi:10.1016/j.molcel.2007.05.001 (2007).
3.Lisitskaya, L., Aravin, A. A. & Kulbachinskiy, A. DNA interference and beyond: structure and functions of prokaryotic Argonaute proteins. Nat Commun 9, 5165, doi:10.1038/s41467-018-07449-7 (2018).
4.Koopal, B. et al. Short prokaryotic Argonaute systems trigger cell death upon detection of invading DNA. Cell 185, 1471-1486 e1419, doi:10.1016/j.cell.2022.03.012 (2022).
產(chǎn)品訂購:sales@amyjet.com
郵政編碼:430070
公司地址:武漢市洪山區(qū)光谷大道35號
光谷總部國際二期時代1棟13樓
提示:本公司所有產(chǎn)品僅供科研使用,不用于臨床診斷。
版權(quán)所有:艾美捷科技有限公司 鄂ICP備10204150號-1 鄂公網(wǎng)安備:42018502004523號
第二類醫(yī)療器械經(jīng)營備案憑證:鄂漢藥監(jiān)械經(jīng)營備20234324號

微信掃碼在線客服