PRODUCT CENTER
特色產(chǎn)品當(dāng)前位置:首頁 > 特色產(chǎn)品
引言
谷氨酸是大腦中最重要的興奮性神經(jīng)遞質(zhì)。許多不同類型的離子型和代謝型受體介導(dǎo)其對神經(jīng)元的刺激信號。但是,就像開關(guān)一樣,“關(guān)閉”信號和“開啟”信號同樣重要。興奮性氨基酸轉(zhuǎn)運體(EAATs)家族通過將谷氨酸回收回神經(jīng)元和星形膠質(zhì)細胞,迅速從突觸間隙和周圍細胞外空間中移除谷氨酸,從而幫助終止谷氨酸的興奮信號。通過這種方式,EAATs管理:
1、維持谷氨酸信號的時間保真度。通過從突觸間隙中移除谷氨酸,它們防止了初始信號發(fā)送后對突觸后受體的過度刺激,這會損害對下一個到達信號的檢測。
2、維持信號的空間保真度,防止谷氨酸從其釋放的突觸中“逃逸”,并刺激意外的突觸外或突觸谷氨酸受體。
3、保護神經(jīng)元免受細胞外空間中過量谷氨酸的毒性作用。谷氨酸受體的加劇或持續(xù)激活啟動了一系列神經(jīng)毒性反應(yīng),最終導(dǎo)致神經(jīng)元功能喪失和細胞死亡,這被稱為興奮毒性。
谷氨酸和陰離子的運輸
EAATs作為共轉(zhuǎn)運蛋白工作,通過將一個K+離子從細胞外運輸出,同時將谷氨酸神經(jīng)遞質(zhì)/分子(也可以運輸天冬氨酸)和三個Na+離子以及一個H+離子帶入細胞內(nèi)(Alleva et al., 2022),見圖1(改編自Freidman et al., 2020)。這種運輸依賴于鈉離子的電化學(xué)梯度,并且由細胞膜上的EAATs的同源或異源三聚體促進(Kovermann et al., 2022)。
EAATs不僅是谷氨酸轉(zhuǎn)運體,還是陰離子通道(Cl+),它們在谷氨酸運輸周期內(nèi)的轉(zhuǎn)換響應(yīng)中打開(Otis & Jahr, 1998)。
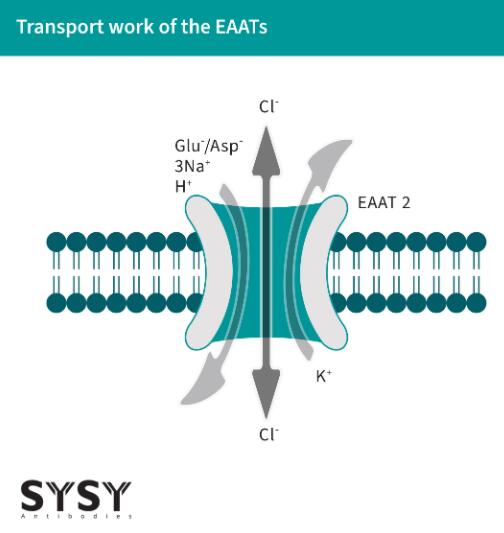
圖1:EAAT對谷氨酸和陰離子的運輸
最近發(fā)表的數(shù)據(jù)表明,EAAT5是EAAT家族中特定于視網(wǎng)膜的成員,它不僅作為谷氨酸的導(dǎo)入器,還作為谷氨酸門控的Cl-通道,特別是在視錐細胞中(Lukasiewcz et al., 2021)。
當(dāng)谷氨酸通過EAATs被吸收進入膠質(zhì)細胞時,它被轉(zhuǎn)化為谷氨酰胺,隨后被運輸回突觸前神經(jīng)元,再轉(zhuǎn)化為谷氨酸,并由VGLUTs的作用被吸收進突觸小泡中。這個過程被稱為谷氨酸-谷氨酰胺循環(huán)(Andersen & Schousboe, 2023),見圖2。
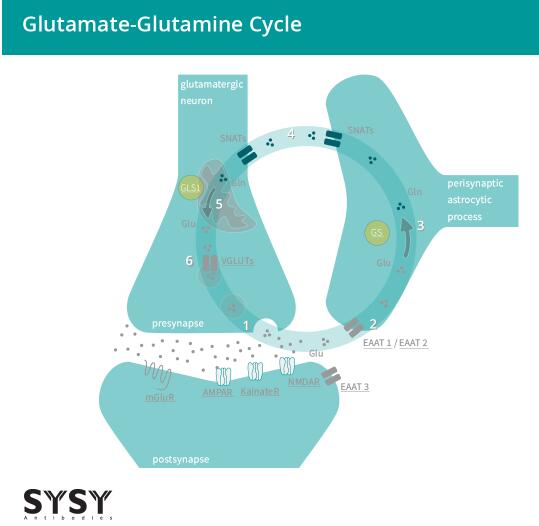
圖2:谷氨酸能神經(jīng)元與星形膠質(zhì)細胞之間的谷氨酸-谷氨酰胺循環(huán)。
(1) 谷氨酸(Glu)被釋放并與離子型和代謝型受體(AMPAR/GluA, KainateR/GluK, NMDAR/GluN, mGluRs)結(jié)合。(2) 谷氨酸主要由星形膠質(zhì)細胞通過興奮性氨基酸轉(zhuǎn)運體EAAT 1/2吸收,部分由神經(jīng)元通過EAAT3吸收。(3) 星形膠質(zhì)細胞的谷氨酰胺合成酶將谷氨酸(Glu)轉(zhuǎn)化為谷氨酰胺(Gln)。(4) 突觸不活躍的谷氨酰胺從星形膠質(zhì)細胞轉(zhuǎn)移到神經(jīng)元。(5) 谷氨酰胺(Gln)通過線粒體谷氨酰胺酶1(GLS1)被轉(zhuǎn)化回谷氨酸(Glu)。(6) 谷氨酸通過囊泡谷氨酸轉(zhuǎn)運體(VGLUTs)被轉(zhuǎn)運進小泡,并準備好進行下一輪的傳遞。
EAAT的分布
EAAT家族目前由5個成員組成,分別是EAAT1到EAAT5,它們在谷氨酸攝取動力學(xué)、氯離子通透性和分布程度上各不相同(Todd & Harding, 2020),見表1。
表1:
protein | gene | molecular mass (mouse) | Cl-conduct. | tissue distribution |
EAAT1 | SLC1A3 | ~ 60 kDa | mod. | Astroglia (low level in lung, spleen, skeletal muscle and testis) |
EAAT2 | SLC1A2 | ~ 62 kDa | low | mainly astroglia; mediates >90% of CNS glutamate reuptake |
EAAT3 | SLC1A1 | ~57 kDa | mod. | all neurons – located on dendrites and axon terminals, epithelial cells of the kidney and the gastrointestinal tract (low level in lung, kidney, skeletal muscle and small intestine) |
EAAT4 | SLC1A6 | ~61 kDa | high | neurons (postsynaptic, dendritic spines) in cerebellum |
EAAT5 | SLC1A7 | ~60 kDa | high | CNS: predominantly retina |
EAAT1:
EAAT1,也稱為GLAST-1,在整個中樞神經(jīng)系統(tǒng)中表達,在小腦的星形膠質(zhì)細胞和Bergmann膠質(zhì)細胞中高度表達。在視網(wǎng)膜中,EAAT1在Müller細胞中表達。
穩(wěn)健的EAAT表達,特別是EAAT1,是成人神經(jīng)干細胞(NSC)表型的廣泛使用的標(biāo)志物(Rieskamp等人,2023)。
我們提供幾種經(jīng)KO驗證的抗體用于檢測EAAT1。
| EAAT1產(chǎn)品 | |||
| 貨號 | 名稱 | 應(yīng)用 | 規(guī)格 |
| 250 103 | EAAT1,rabbit,polyclonal,affinity purified K.O. extracellulardomain | WB | 50 μg |
| 250 113 | EAAT1,rabbit,polyclonal,affinity purified K.O. cytoplasmic domain | WB IP ICC IHC | 50 μg |
| 250 114 | EAAT1,Guinea pig,polyclonal,antiserum cytoplasmic domain | WB ICC IHC IHC-P | 100μl |
| 250 116 | EAAT1,chicken,polyclonal,IgY fraction cytoplasmic domain | WB ICC IHC | 200 μl |
| 250-11P | EAAT1,control peptide cytoplasmic domain | 100 μg | |
| 250-1P | EAAT1,control peptide extracellulardomain | 100 μg | |
|
|
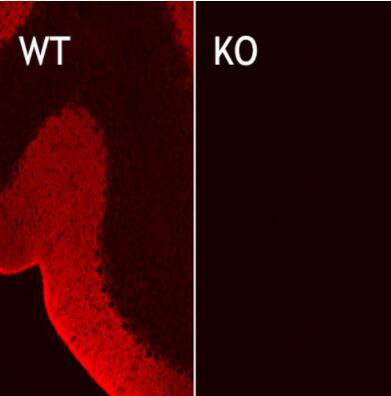
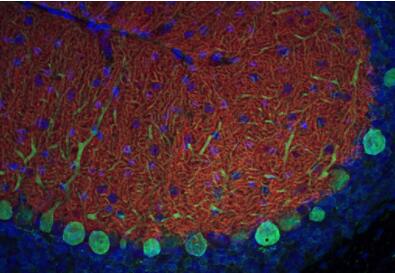
圖3:用兔多克隆抗EAAT1(目錄號250 113,稀釋度1:5000;紅色)對野生型(WT)和敲除型(KO)動物的小鼠小腦中的EAAT1進行間接免疫染色。圖片提供:奧斯陸大學(xué)基礎(chǔ)醫(yī)學(xué)研究所解剖學(xué)系周云和Niels Christian Danbolt | 圖4:PFA固定的小鼠小腦用豚鼠多克隆抗EAAT1(目錄號250 114,稀釋度1:500;紅色)和兔抗細小白蛋白(目錄號195 002,稀釋度1:100;綠色)進行間接免疫染色。DAPI染色(藍色)顯示細胞核。 |
EAAT2:
EAAT2,也稱為GLT-1,是大腦中EAATs中含量最豐富的。它主要位于星形膠質(zhì)細胞分支,分別在小腦和海馬體中高度表達(Yeung等人,2021)。最近,Dahlmanns等人在2023年綜述了幾種神經(jīng)退行性疾病,如阿爾茨海默病、多發(fā)性硬化癥和肌萎縮側(cè)索硬化癥(ALS),發(fā)現(xiàn)EAAT2減少。在此背景下,值得注意的是,γ分泌酶的活性亞基早老素1(PS1)直接與EAAT2相互作用,并影響轉(zhuǎn)運蛋白的細胞表面定位(Perrin等人,2024)。
對于EAAT2的檢測,我們可以提供優(yōu)秀的、KO驗證的、多克隆的兔和豚鼠抗體。此外,還有一種小鼠單克隆抗體,在WB、ICC、IHC和FFPE(IHC-P)應(yīng)用中顯示出優(yōu)異的結(jié)果。
| EAAT2產(chǎn)品 | |||
| 貨號 | 名稱 | 應(yīng)用 | 規(guī)格 |
| 250 203 | EAAT2,rabbit,polyclonal,affinity purified (K.O. extracellular domain | WB IHC IHC-P | 50 μg |
| 250 204 | EAAT2,Guineapig,polyclonal,antiserum (K.O. extracellular domain | WB ICC IHC IHC-P | 100 μl |
| 250 211 | EAAT2,mouse,monoclonal,purified IgG extracellular domain | WB ICC IHCIHC-P | 100 μg |
| 250-2P | EAAT2,control peptide extracellular domain | 100 μg | |
|
|
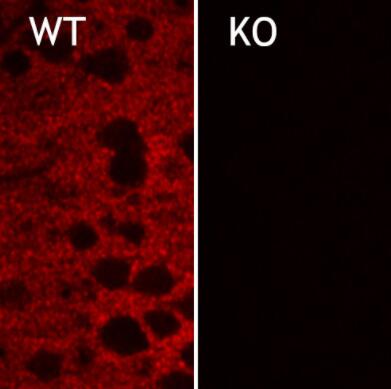
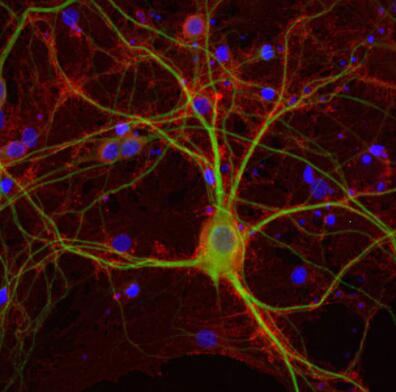
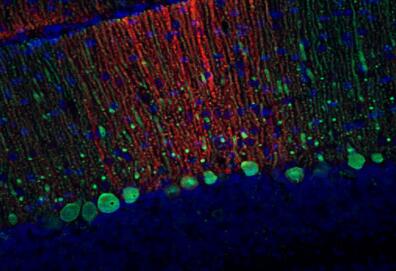
圖5:雜合子(+/-)和敲除(-/-)小鼠新皮層中EAAT2的間接免疫染色(分類號250 203,稀釋度1:2000;紅色)。圖片提供:奧斯陸大學(xué)基礎(chǔ)醫(yī)學(xué)研究所解剖學(xué)系周云和Niels Christian Danbolt | 圖6:PFA固定的大鼠海馬神經(jīng)元用抗EAAT2(目錄號250 204,稀釋度1:500;紅色)和小鼠抗MAP2(目錄編號188 011,稀釋度1:100;綠色)進行間接免疫染色。DAPI染色(藍色)顯示細胞核。 |
EAAT3:
EAAT3,也稱為EAAC1或SLC1A1,是大腦中的“神經(jīng)元”谷氨酸轉(zhuǎn)運蛋白,主要位于軸突末端和大腦皮層、海馬、紋狀體和基底節(jié)的樹突(Escobar等人,2019)。最新的研究結(jié)果假設(shè)EAAT3表達與強迫癥(OCD)之間存在聯(lián)系,Escobar等人于2019年對此進行了綜述。中樞神經(jīng)系統(tǒng)外的EAAT3可以在肺、小腸、骨骼肌、腎外髓質(zhì)、髓質(zhì)射線和皮質(zhì)中發(fā)現(xiàn)(Todd&Hardingham,2020)。
我們的產(chǎn)品組合中提供2種優(yōu)秀的多克隆兔抗體。
| EAAT3產(chǎn)品 | |||
| 貨號 | 名稱 | 應(yīng)用 | 規(guī)格 |
| 250 303 | EAAT3,rabbit,polyclonal,affinity purified currently not available | ICC IHC | 50 μg |
| cytoplasmic domain | |||
| 250 313 | EAAT3,rabbit,polyclonal,affinity purified cytoplasmic domain | WB | 50 μg |
| 250-31P | EAAT3,control peptide cytoplasmic domain | 100 μg | |
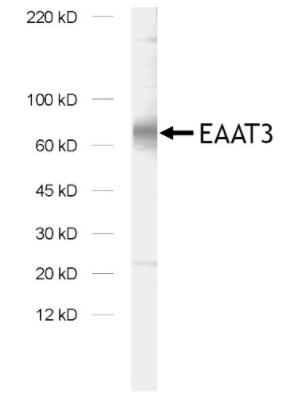
圖7:用兔多克隆抗EAAT3(目錄號250 313)對大鼠腦突觸膜部分(LP1)中的EAAT3進行免疫印跡。
EAAT4:
EAAT4,也稱為SLC1A6,是一種主要的神經(jīng)元谷氨酸進口商。它主要位于小腦的浦肯野細胞上,在前腦和中腦的某些亞區(qū)有一些稀疏的表達(Massie等人,2008)。最近的數(shù)據(jù)顯示,EAAT4的表達遵循類似于醛縮酶C(zebrin)表達的副矢狀帶模式,產(chǎn)生了EAAT4水平高低的分子多樣性浦肯野細胞的微區(qū)(Malhotra等人,2021)。與EAAT1和EAAT2相比,EAAT4的谷氨酸轉(zhuǎn)運能力相當(dāng)?shù)停虼艘恍┭芯咳藛T推測EAAT4在生理上主要作為陰離子通道發(fā)揮作用(Suslova等人,2023)。
我們的產(chǎn)品組合中新增了2種針對EAAT4的優(yōu)秀兔多克隆抗體。
|
|
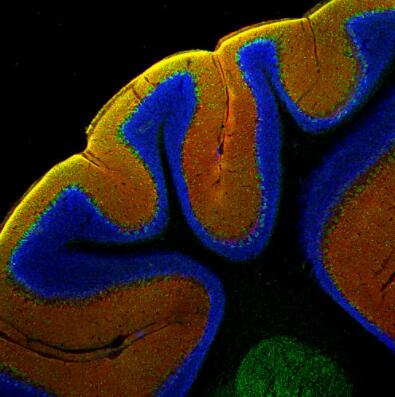
圖8:甲醛固定的小鼠小腦切片(矢狀)兔抗EAAT4抗體(目錄號250 413,稀釋度1:500,紅色)和豚鼠抗Calbindin抗體(目錄編號214 318,稀釋度1:100,綠色)的間接免疫染色。DAPI染色(藍色)顯示細胞核。 | 圖9:甲醛固定的小鼠小腦切片(冠狀)兔抗EAAT4抗體(目錄號250 413,稀釋度1:500,紅色)和豚鼠抗Calbindin抗體(目錄編號214 318,稀釋度1:100,綠色)的間接免疫染色。DAPI染色(藍色)顯示細胞核。 |
EAAT5:
EAAT5蛋白,有時也稱為AAAT,主要存在于脊椎動物視網(wǎng)膜中,在那里它(與EAAT2一起)將谷氨酸轉(zhuǎn)運到視桿細胞、視錐細胞和視桿雙極細胞中。EAAT5轉(zhuǎn)運蛋白聚集在突觸前桿和錐活性區(qū)的突觸帶下方,位置完美,可以捕獲最近釋放的谷氨酸。對EAAT5的抑制表明,該蛋白對視網(wǎng)膜中甘氨酸能無長突細胞(AII無長突細胞)的時間信號分辨很重要(Tang等人,2022)。在中樞神經(jīng)系統(tǒng)外,EAAT5也在肝臟、腎臟、腸道、心臟、肺和肌肉中表達。我們提供了一種針對EAAT5的多克隆豚鼠KO驗證抗體。
| EAAT5產(chǎn)品 | |||
| 貨號 | 名稱 | 應(yīng)用 | 規(guī)格 |
| 250 504 | EAAT5, Guinea pig, polyclonal, antiserum | IHC IHC-P | 100 ?l |
|
|
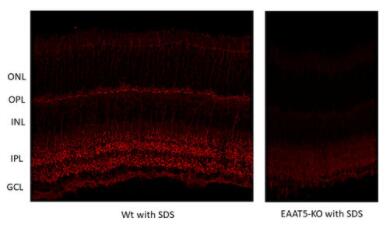
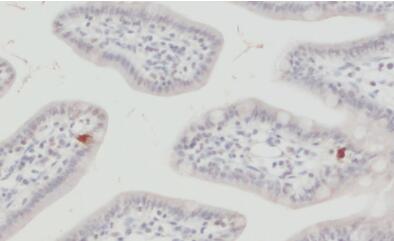
圖10:野生型(WT)和敲除型(KO)動物小鼠視網(wǎng)膜中EAAT5的間接免疫染色(分類號250 504,稀釋度1:2000;紅色)。根據(jù)(Gehlen等人,2021),用4%甲醛浸泡固定組織,用1%SDS進行抗原修復(fù)。圖片提供:Christoph Aretzweiler von Schwartzenberg和Frank Müller,德國Forschungszentrum Jülich生物信息處理、分子和細胞生理學(xué)研究所(IBI-1) | 圖11:用豚鼠抗EAAT5抗體(目錄號250 504,稀釋1:1000,DAB;棕色)對福爾馬林固定石蠟包埋(FFPE)小鼠回腸切片進行間接免疫染色。蘇木精染色(藍色)顯示細胞核。 |
產(chǎn)品:
| 貨號 | 名稱 | 應(yīng)用 | 規(guī)格 |
| 250 103 | EAAT1,rabbit,polyclonal,affinity purified(K.0. extracellular domain | WB | 50 μg |
| 250 113 | EAAT1,rabbit,polyclonal,affinity purified(K.O.) cytoplasmic domain | WB IP ICC IHC | 50 μg |
| 250 114 | EAAT1,Guineapig,polyclonal,antiserum cytoplasmic domain | WB ICC IHCIHC-P | 100 μl |
| 250 116 | EAAT1,chicken,polyclonal,IgY fraction cytoplasmicdomain | WB ICC IHC | 200μl |
| 250-11P | EAAT1,control peptide cytoplasmicdomain | 100pg | |
| 250-1P | EAAT1,control peptide extracellular domain | 100 μg | |
| 250 203 | EAAT2,rabbit,polyclonal,affinity purified(K.O.) extracellulardomain | WB IHC IHC-P | 50 μg |
| 250 204 | EAAT2,Guinea pig,polyclonal,antiserum (K.0. extracellular domain | WB ICC IHC IHC-P | 100 μl |
| 250 211 | EAAT2,mouse,monoclonal,purified IgG extracellular domain | WB ICC IHCIHC-P | 100pg |
| 250-2P | EAAT2,control peptide extracellular domain | 100 μg | |
| 250 303 | EAAT3,rabbit,polyclonal,affinity purified currently not available cytoplasmic domain | 50 μg | |
| CC IHC | |||
| 250313 | EAAT3,rabbit,polyclonal,affinity purified cytoplasmic domain | WB | 50 pg |
| 250-31P | EAAT3,control peptide cytoplasmicdomain | 100 μg | |
| 250 403 | EAAT4,rabbit,polyclonal,affinity purified | WB | 50 μg |
| 250 413 | EAAT4, rabbit, polyclonal, affinity purified | WB ICC IHC IHC-P | 50 ?g |
| 250 504 | EAAT5, Guinea pig, polyclonal, antiserum | IHC IHC-P | 100 ?l |
文獻參考:
Alleva et al., 2022: Molecular Basis of Coupled Transport and Anion Conduction in Excitatory Amino Acid Transporters. PMID: 33587237
Andersen & Schousboe, 2023: Glial Glutamine Homeostasis in Health and Disease. PMID: 36322369
Dahlmanns et al., 2023: Glial Glutamate Transporter-Mediated Plasticity: System xc-/xCT/SLC7A11 and EAAT1/2 in Brain Diseases. PMID: 37005761
Escobar et al., 2019: The Neuronal Glutamate Transporter EAAT3 in Obsessive-Compulsive Disorder. PMID: 31803055
Freidman et al., 2020: Amino Acid Transporters and Exchangers from the SLC1A Family: Structure, Mechanism and Roles in Physiology and Cancer. PMID: 31981058
Kovermann et al., 2022: Cellular Physiology and Pathophysiology of EAAT Anion Channels. PMID: 35087380
Lukasiewcz et al., 2021: EAAT5 Glutamate Transporter-Mediated Inhibition in the Vertebrate Retina. PMID: 34025361
Malhotra et al., 2021: Climbing Fiber-Mediated Spillover Transmission to Interneurons Is Regulated by EAAT4. PMID: 34400517
Massie et al., 2008: High-affinity Na+/K+-dependent glutamate transporter EAAT4 is expressed throughout the rat fore- and midbrain. PMID: 18770868
Otis & Jahr, 1998: Anion currents and predicted glutamate flux through a neuronal glutamate transporter. PMID: 9736633
Perrin et al., 2024: Identification of PS1/gamma-secretase and glutamate transporter GLT-1 interaction site. PMID: 38499151
Rieskamp et al., 2023: Excitatory amino acid transporter 1 supports adult hippocampal neural stem cell self-renewal. PMID: 37534178
Suslova et al., 2023: Apo state pore opening as functional basis of increased EAAT anion channel activity in episodic ataxia 6. PMID: 37538371
Tang et al., 2022: Glutamate Transporters EAAT2 and EAAT5 Differentially Shape Synaptic Transmission from Rod Bipolar Cell Terminals. PMID: 35523583
Todd & Hardingham, 2020: The Regulation of Astrocytic Glutamate Transporters in Health and Neurodegenerative Diseases. PMID: 33348528
Yeung et al., 2021: EAAT2 Expression in the Hippocampus, Subiculum, Entorhinal Cortex and Superior Temporal Gyrus in Alzheimer’s Disease. PMID: 34588956
產(chǎn)品訂購:sales@amyjet.com
郵政編碼:430070
公司地址:武漢市洪山區(qū)光谷大道35號
光谷總部國際二期時代1棟13樓
提示:本公司所有產(chǎn)品僅供科研使用,不用于臨床診斷。
版權(quán)所有:艾美捷科技有限公司 鄂ICP備10204150號-1 鄂公網(wǎng)安備:42018502004523號
第二類醫(yī)療器械經(jīng)營備案憑證:鄂漢藥監(jiān)械經(jīng)營備20234324號

微信掃碼在線客服