PRODUCT CENTER
特色產(chǎn)品當前位置:首頁 > 特色產(chǎn)品
【前情提示】:Flipper-TR® :一種革命性的新型活細胞膜張力熒光探針,點擊了解
細胞膜張力調(diào)節(jié)許多細胞過程,如胞吞,胞吐,分裂和細胞運動1-6,所有這些過程都需要重塑細胞形態(tài),而這反過來又依賴于細胞膜變形和肌動蛋白細胞骨架重塑之間的相互作用1,3,5-9 , 本通訊主要討論細胞運動相關的形態(tài)發(fā)生過程中Actin細胞骨架與膜張力之間的協(xié)同作用。

細胞定向運動需要在細胞的前緣和后緣進行Actin細胞骨架的動態(tài)重組,其中前緣的肌動蛋白的片狀脂質(zhì)突起與后緣的回縮平行地向前或向后拉動細胞定向運動,同時伴隨著粘附位點的改變和RhoA介導的肌動蛋白收縮8,10,11。由于運動細胞前緣受Actin突起的推動,后緣發(fā)生收縮,細胞膜發(fā)生變形,膜張力發(fā)生變化,定向運動依賴于Actin細胞骨架重塑與變形引起的膜張力變化之間的平衡(圖1)。
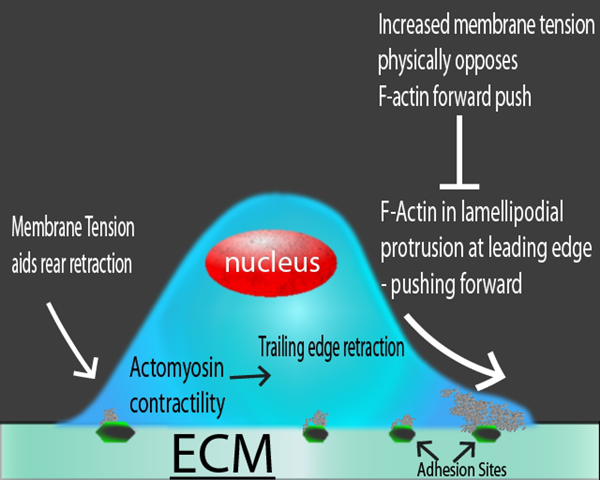
圖1:遷移細胞中不同作用力的示意圖。在前緣的基于肌動蛋白的突起(即片狀脂膜)將細胞向前推動,而膜張力在物理上抵抗這種運動。在后緣,膜張力將幫助Actin介導的收縮。
膜張力和Actin細胞骨架的動態(tài)重組是如何相互作用的了?上圖中細胞形態(tài)變化的運動模型是從早期2D細胞運動實驗中獲得的,該實驗是用魚角膜細胞的片狀脂質(zhì)體片段或活的完整角膜細胞進行的8,12。該模型假設,運動細胞前緣的聚合Actin網(wǎng)絡從內(nèi)部推動細胞膜,產(chǎn)生與肌動蛋白突起相對的膜張力,在聚合的肌動蛋白網(wǎng)絡上快速平衡并施加全局恒定力。
在前緣的中心,高密度的肌動蛋白纖維導致每根絲的膜張力降低,從而使肌動蛋白纖維迅速聚合并驅(qū)動前緣向外伸出。隨著細胞側面肌纖維密度減小,每根肌纖維的阻力增加,直到膜張力負荷使肌動蛋白聚合停止,從而形成了細胞的前角。類似地,早期對成纖維細胞的研究也發(fā)現(xiàn),膜張力和突起數(shù)呈負相關1。
同時,遷移細胞的后緣有一個可分解的肌動蛋白網(wǎng)絡,膜張力的增加會使其解聚加速,從而導致后緣收縮12-14。運動性角膜細胞的活細胞成像研究表明,平面張力(膜本身的張力)和膜細胞骨架附著張力(粘連和肌動球蛋白介導的收縮力)均調(diào)節(jié)運動細胞的膜張力15,實際上總膜張力由兩個因素決定8。Lieber等還證實,隨著前緣肌動蛋白聚合的增加,肌纖維向前的推進力也更強從而導致更高的膜張力,反之亦然,在后緣,膜張力與肌球蛋白收縮水平呈負相關,與粘附強度呈正相關15。
最近,在成纖維細胞和成纖維樣細胞中的研究進一步闡明了Actin骨架與膜張力之間的相互作用。對成纖維細胞進行的高分辨率顯微鏡研究顯示16,在片狀脂質(zhì)體突起形成早期,當膜貯液充足時,突起的長度就很明顯,而膜張力卻很低,隨著片狀脂膜在膜上向外推,膜張力增加,延伸的突起使質(zhì)膜上的褶皺和曲線平滑,膜貯液耗盡,膜張力迅速增加,同時肌動蛋白細胞骨架重組,導致突起長度縮短16。
在嗜中性粒細胞中,遷移細胞前緣突起的增加與膜張力的增加和除前緣外的其他細胞位置突起的減少是一致的。另一項成纖維細胞研究揭示了膜張力的周期性振蕩與片狀脂質(zhì)體突起形態(tài)呈負相關17。當膜張力增加時,突起的尺寸(寬度減小)和形狀(向上彎曲)都會發(fā)生相應變化,并且隨著張力的消失,突起會恢復正常的大小,形狀和生長活動17。肌動蛋白相關蛋白FBP17(Formin-binding protein)是一種膜彎曲蛋白,是WASP / N-WASP依賴型肌動蛋白成核的活化劑,位于COS-1(成纖維樣細胞)細胞遷移的前緣。膜張力通過觸發(fā)FBP17的膜分離來抑制FBP17的功能,膜張力抑制肌動蛋白的聚合反應從而提供了一個信號傳遞途徑18,19,這表明存在負反饋回路:增加的膜張力通過使肌動蛋白組裝分子失活而進一步抑制突起生成8,18,19。其他Actin結合蛋白和MAP蛋白也會參與調(diào)節(jié)前緣突起的肌動蛋白網(wǎng)絡的動態(tài)重組20。
摘要
膜張力的功能相關性超出了生物物理范疇,因為膜張力以及其他膜屬性,如順序和電位幾乎影響著任何一個重要的生物過程。測量活細胞膜的張力和動態(tài)變化是一項非常困難的工作,挑戰(zhàn)包括在三維細胞培養(yǎng)中驗證在二維細胞培養(yǎng)獲得的結果,剖析在不同細胞類型之間膜張力的調(diào)節(jié)變化,以及不同細胞狀態(tài)之間膜張力的變化8。
好消息是經(jīng)過科研工作者們不懈的努力,目前市面上已經(jīng)有專門用于實時檢測活細胞中膜張力變化的械敏感的熒光機探針了9,22,23。艾美捷科技為您推薦Cytoskeleton生產(chǎn)的一種新型的熒光探針,以幫助生物物理學家和細胞生物學家更深入地了解膜張力如何與細胞骨架相互作用以調(diào)節(jié)基本的細胞功能的。
艾美捷科技為您提供Cytoskeleton的Flipper-TR®活細胞膜張力檢測探針 :
| 貨號 | 產(chǎn)品名稱 | 規(guī)格 |
| CY-SC020 | Flipper-TR Membrane Tension Probe | 50 nmol |
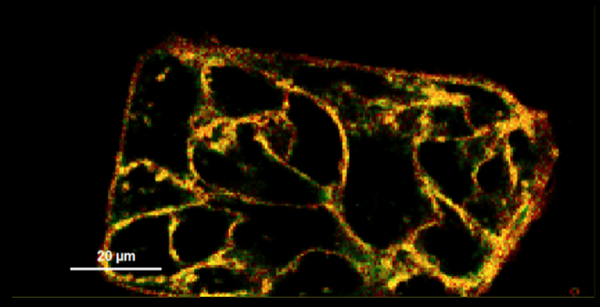
圖2:用Flipper-TR膜張力探針(目錄號CY-SC020)染色的細胞膜的熒光壽命圖。綠色表示中等張力,黃色/橙色/紅色表示較高的張力,藍色表示較低的張力。圖片由Colom等提供, 2018.
此外,Cytoskeleton還提供用于F-actin,微管,DNA和溶酶體的活細胞成像探針,以及用于研究細胞中G-actin/F-actin水平和tubulin/微管以及體外相互作用的生化檢測試劑盒 。
活細胞成像試劑盒:
| 產(chǎn)品名稱 | 貨號 |
| SiR-Actin Kit | (Cat. # CY-SC001) |
| SiR-Tubulin Kit | (Cat. # CY-SC002) |
| Cytoskeleton Kit (包括SiR-Actin, SiR-Tubulin,和Verapamil) | (Cat. # CY-SC006) |
| SiR-DNA Kit | (Cat. # CY-SC007) |
| SiR-Lysosome Kit | (Cat. # CY-SC012) |
| SiR700-Actin Kit | (Cat. # CY-SC013) |
| SiR700-Tubulin Kit | (Cat. # CY-SC014) |
| SiR700-DNA Kit | (Cat. # CY-SC015) |
| SiR700-Lysosome Kit | (Cat. # CY-SC016) |
Actin生化試劑盒:
| 產(chǎn)品名稱 | 貨號 |
| 肌動蛋白結合蛋白 Spin-Down生化檢測試劑盒(兔骨骼肌actin) | (Cat. # BK001) |
| 肌動蛋白結合蛋白 Spin-Down生化檢測試劑盒(人血小板actin) | (Cat. # BK013) |
| Actin聚合生化檢測試劑盒(熒光法:兔骨骼肌actin) | (Cat. # BK003) |
| G-Actin/F-actin In Vivo生化檢測試劑盒 | (Cat. # BK037) |
Tubulin生化試劑盒:
| 產(chǎn)品名稱 | 貨號 |
| Tubulin聚合生化檢測試劑盒(比色法) | (Cat. # BK006P) |
| Tubulin聚合生化檢測試劑盒(熒光法) | (Cat. # BK011P) |
| 微管結合蛋白 Spin-Down生化檢測試劑盒 | (Cat. # BK029) |
| 微管/Tubulin In Vivo生化檢測試劑盒 | (Cat. # BK038) |
參考文獻:
Raucher D. and Sheetz M.P. 2000. Cell spreading and lamellipodial extension rate is regulated by membrane tension. J. Cell Biol. 148, 127-136.
Keren K. 2011. Cell motility: the integrating role of the plasma membrane. Eur. Biophys. J. 40, 1013-1027.
Diz-Munoz A. et al. 2013. Use the force: Membrane tension as an organizer of cell shape and motility. Trends Cell Biol. 23, 47-53.
Masters T.A. et al. 2013. Plasma membrane tension orchestrates membrane trafficking, cytoskeletal remodeling, and biochemical signaling during phagocytosis. Proc. Natl. Acad. Sci. U.S.A. 110, 11875-11880.
Clark A.G. et al. 2014. Stresses at the cell surface during animal cell morphogenesis. Curr. Biol. 24, R484-R494.
Sens P. and Plastino J. 2015. Membrane tension and cytoskeleton organization in cell motility. J. Phys. Condens. Matter. 27, 273103.
Houk A.R. et al. 2012. Membrane tension maintains cell polarity by confining signals to the leading edge during neutrophil migration. Cell. 148, 175-188.
Pontes B. et al. 2017. Membrane tension: A challenging but universal physical parameter in cell biology. Semin. Cell Dev. Biol. 71, 30-41.
Colom A. et al. 2018. A fluorescent membrane tension probe. Nat. Chem. 10, 1118-1125.
Warner H. et al. 2019. Control of adhesion and protrusion in cell migration by Rho GTPases. Curr. Opin. Cell Biol.56, 64-70.
Lawson C.D. and Ridley A.J. 2018. Rho GTPase signaling complexes in cell migration and invasion. J. Cell Biol. 217, 447-457.
Keren K.et al. 2008. Mechanism of shape determination in motile cells. Nature. 453, 475–480.
Kozlov M.M. and Mogilner A. 2007. Model of polarization and bistability of cell fragments. Biophys. J. 93, 3811–3819.
Ofer N. et al. 2011. Actin disassembly clock determines shape and speed of lamellipodial fragments. Proc. Natl. Acad. Sci. U.S.A. 108, 20394–20399.
Lieber A.D. et al. 2013. Membrane tension in rapidly moving cells is determined by cytoskeletal forces. Curr. Biol.23, 1409-1417.
Gauthier N.C. et al. 2011. Temporary increase in plasma membrane tension coordinates the activation of exocytosis and contraction during cell spreading. Proc. Natl. Acad. Sci. U.S.A. 108, 14467-14472.
Pontes B. et al. 2017. Membrane tension controls adhesion positioning at the leading edge of cells. J. Cell Biol. 216, 2959-2977.
Zegers M.M. and Friedl P. 2015. membrane tension into cytoskeletal action by FBP17. Dev. Cell. 33, 628-630.
Tsujita K. et al. 2015. Feedback regulation between plasma membrane tension and membrane-bending proteins organizes cell polarity during leading edge formation. Nat. Cell Biol. 17, 749-758.
Simon C. et al. 2018. Interplay between membrane tension and the actin cytoskeleton determines shape changes. Phys. Biol. 15, 065004.
Diz-Munoz A. et al. 2016. Membrane tension acts through PLD2 and mTORC2 to limit actin network assembly during neutrophil migration. PLoS Biol. 14, e1002474.
Molin M.D. et al. 2015. Fluorescent flippers for mechanosensitive membrane probes. J. Am. Chem. Soc. 137, 568-571.
Goujon A. et al. 2019. Mechanosensitive fluorescent probes to image membrane tension in mitochondria, endoplasmic reticulum, and lysosomes. J. Am. Chem. Soc. 141, 3380-3384.
Cytoskeleton公司成立于1993年,專注于生物化學和細胞過程研究中的純化蛋白和便捷試劑盒開發(fā)與生產(chǎn)。公司提供藥物篩選、信號轉(zhuǎn)導、蛋白質(zhì)轉(zhuǎn)錄后修飾(PTM)、細胞骨架研究相關的系列試劑盒和產(chǎn)品,尤其以細胞骨架相關研究見長,既能滿足于樣品較少的科學研究,也可以用于小規(guī)模篩選研究和高通量大規(guī)模篩選研究。此外,公司還提供微管蛋白,肌動蛋白,小G蛋白,GAPs,GEFs等現(xiàn)有產(chǎn)品的藥物篩選服務。
作為Cytoskeleton在中國的區(qū)域總代理,艾美捷科技有限公司將為中國客戶提供最全面的Cytoskeleton產(chǎn)品與服務。
產(chǎn)品訂購:sales@amyjet.com
郵政編碼:430070
公司地址:武漢市洪山區(qū)光谷大道35號
光谷總部國際二期時代1棟13樓
提示:本公司所有產(chǎn)品僅供科研使用,不用于臨床診斷。
版權所有:艾美捷科技有限公司 鄂ICP備10204150號-1 鄂公網(wǎng)安備:42018502004523號
第二類醫(yī)療器械經(jīng)營備案憑證:鄂漢藥監(jiān)械經(jīng)營備20234324號

微信掃碼在線客服